Fleurs d’Iris : un tunnel pour pollinisateurs
Iris

Le genre Iris se démarque, y compris au sein de sa propre famille, par une architecture florale très particulière ; chaque fleur est en fait formée de trois « sous-fleurs » réunies en une avec un style à trois branches qui prend des allures de corolle. On se doute bien qu’une structure aussi complexe et originale a du se mettre en place au cours de l’évolution sous la pression de la sélection liée à la pollinisation par les insectes. Dans la chronique Fascinante orchidée du Nord nous avons exploré tous les secrets de cette fleur et comment en interpréter les différentes parties, étape indispensable si on veut comprendre ce qui se passe lors d’une visite d’insecte pollinisateur. Quel est notamment le rôle des trois cercles (tépales externes, styles pétaloïdes et tépales internes) qui structurent la fleur ? Quels avantages une telle architecture confère t’elle en termes de succès reproductif ?
Un tube au bout du tunnel

Appréhender la structure interne en 3D d’une fleur d’un grand iris horticole classique (ceux que l’on voit partout) n’est pas chose facile bien qu’elle soit de taille très conséquente ; avec les trois cercles de « pétales » colorés dressés, étalés ou rabattus, dotés de franges plissés pour certains, la vue ne porte pas loin. Il faut soit se résoudre à casser ou arracher une ou plusieurs des pièces ou observer d’autres espèces aux fleurs moins opulentes et moins chargées en fioritures comme l’iris faux-acore des fossés, canaux et bords d’étangs, l’espèce sauvage la plus commune en France. Avec elle, on réussit à voir « à travers » la fleur en se plaçant à sa hauteur de profil. On constate alors que chaque style pétaloïde qui coiffe une étamine plaquée et cachée forme avec la languette rétrécie (onglet) du tépale externe situé en-dessous un tunnel étroit, gracieusement courbé, qui glisse en pente douce vers le cœur de la fleur : un tunnel avec les côtés ouverts (mais presque pas dans d’autres espèces où style et tépale se touchent).
-

- Fleur d’iris faux-acore avec ses trois unités

-

- La branche pétaloïde du style bombée laisse un large espace au-dessus du tépale externe mais se rabat dessus au bout

-

- Juste sous le style se trouve l’anthère ouverte très proche du tépale en dessous ; en fin de floraison, l’allongement du filet amène l’anthère hors de l’entrée

Pour voir la suite, il faut couper le tépale à sa base et là, on tombe sur une colonne centrale dressée, la hampe du style qui porte les trois branches déployées au-dessus, portée elle-même sur l’ovaire. A ce niveau, le tunnel se rétrécit fortement et s’incurve brusquement à angle droit pour plonger le long de la hampe du style, formant un court tube très étroit et vertical. Vu de dessus sur une fleur découpée, on voit trois trous qui correspondent aux trois tubes, invisibles de l’extérieur, et qui encadrent la base du style branchu. La fleur d’iris combine donc une structure à deux lèvres avec tunnel et tube.
-

- Si on rabat le tépale externe, on découvre le style déployé au dessus et qui sous-tend étroitement une étamine.

-

- Fleur d’Iris de Hollande : style et tépale sont presque jointifs et, de profil, on ne voit pas l’intérieur du tunnel

-

- Etamine collée contre le style : l’anthère longue s’ouvre latéralement pour libérer le pollen.

L’appât du nectar
-
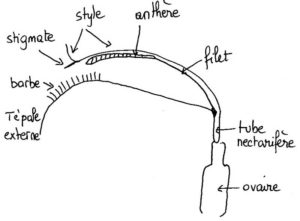
- Coupe dans une fleur d’iris montrant la position du tube à nectar.
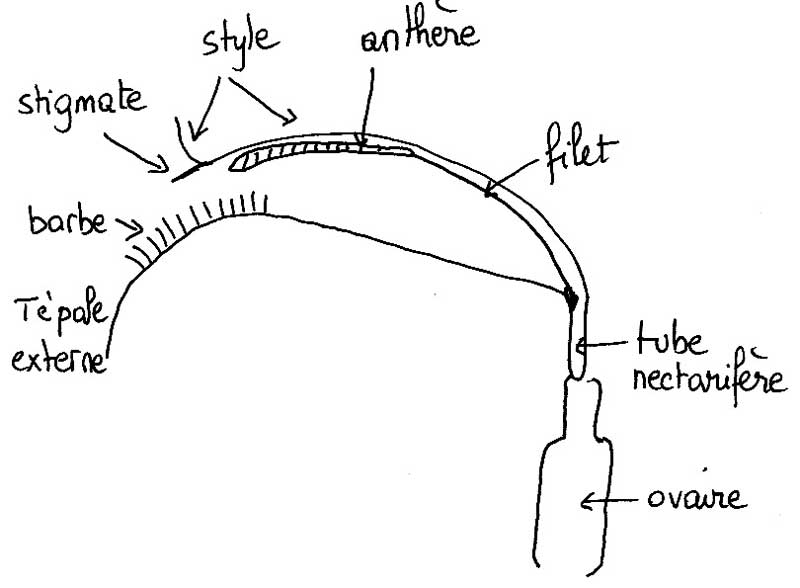
Toute fleur pollinisée par les insectes (entomophile), pour avoir des chances d’être visitée, doit offrir (ou plus subtilement parfois faire semblant) une récompense sonnante et trébuchante sous forme de substance utile à l’insecte ; le plus souvent c’est de la nourriture. Les fleurs d’Iris produisent un nectar abondant secrété par un nectaire glandulaire. Cet organe diffus se trouve justement à la base des tépales externes (les labelles rabattus au premier plan), côté intérieur, autrement dit bien caché au fond du tube que nous venons de repérer. Selon les espèces, il peut être strictement localisé sur la base du tépale externe ou s’étendre plus ou moins autour de la base du style ou aller jusqu’à la base des étamines, voire même jusqu’à celle des tépales internes (le troisième rang dressé au centre de la fleur).
-

- Fleur d’Iris de hollande sur laquelle on a enlevé les deux cercles de tépales : il ne reste que les trois branches du style et les étamines au sommet de l’ovaire

-

- A la base de la fourche du style, on trouve trois ouvertures qui correspondent aux tubes courts qui s’enfoncent dans la fleur à la verticale et où se trouve le nectar.

-

- Sur cette fleur, on a soulevé le style et, en vue plongeante, on aperçoit au fond l’entrée de deux des trois tubes aux angles de l’axe du style.

Là encore, les iris et les autres genres proches de la tribu des iridoïdés se démarquent du reste de la famille des Iridacées : chez tous les autres, les nectaires se trouvent non pas à la base des tépales mais dans les parois de l’ovaire, vers la base (nectaires septaux) ; cette localisation qui se retrouve chez de nombreuses autres monocotylédones semble bien être ancestrale; donc, les nectaires au fond du tube des tépales sont une innovation des iris et des genres alliés. Une telle innovation ne peut qu’être directement liée à la pollinisation : le visiteur intéressé par le nectar devra aller le chercher au fond de ce tube floral étroit, qui fait un coude avec le reste de la fleur et auquel on accède par un tunnel !

Bourdon visitant une fleur d’iris faux-acore
Guider les visiteurs

La gamme des pollinisateurs des iris en général reste assez étroite et se cantonne aux abeilles et bourdons essentiellement ; on est loin de la diversité observée dans d’autres genres de la famille des Iridacées comme les glaïeuls où d’une part la gamme d’insectes est bien plus large avec des papillons de jour ou de nuit, des mouches à très longue trompe, des coléoptères mais aussi des oiseaux (dont les souï-mangas, l’équivalent écologique des colibris en terre africaine). Ceci peut sans doute être mis en relation avec la quasi-absence de signaux olfactifs pour l’attraction des pollinisateurs : chez les iris, pas de parfum envoûtant comme celui des glaïeuls d’Abyssinie par exemple. Primauté aux signaux visuels et tactiles avec les ornementations très variées des tépales externes largement étalés et déployés, les pièces maîtresses dans la fleur d’iris. La diversité est au rendez-vous pour ces signaux « guides à nectar » qui orientent les visiteurs vers le tunnel et le tube : nervures saillantes ; tapis de « barbe » formée de poils multicellulaires très durs formant une brosse ; dessins variés de taches et de motifs ; palette de couleurs ; chez certains, une crête formant une émergence centrale en long ou des appendices latéraux plus ou moins sinueux.
-

- Fleur d’iris d’Allemagne : le style a été enlevé ce qui révèle l’étamine qui surplombe la piste barbue du tépale externe, la piste d’atterrissage.

-

- Vue de face, l’entrée reste étroite entre le style rabattu et le tépale barbu avec son bombement central blanc et ses nervures violacées

-

- Fleur d’iris faux-acore vue par en dessous : les étamines deviennent alors visibles sous les styles

-

- Dessin coloré qui jalonne l’entrée vers le coeur de la fleur

Selon les espèces, l’un ou l’autre ou plusieurs de ces dispositifs se trouvent réunis et témoignent de l’évolution qui a explosé au sein de ce genre fort de 270 espèces. Cette évolution semble d’ailleurs encore plus foisonnante qu’il n’y paraît : ainsi, dans le cas des iris à crête (voir ci-dessus), soit 21% des espèces connues, on a pu montrer (1 et 2) que cet organe, la crête, était apparu au moins 5 fois indépendamment dans le genre Iris, témoin de la richesse de diversification à l’œuvre et de la pression de sélection imposée par la pollinisation.
-

- La brosse de poils dressés des iris barbus rehausse le tépale externe et oblige le pollinisateur à toucher l’anthère sous le style.

La route du nectar

La forme rabattue des tépales externes associée à leur consistance souvent renforcée par des nervures et des excroissances en fait une piste d’atterrissage idéale : l’insecte a juste à choisir par laquelle des trois pistes il va atterrir ! Là, le chemin est tracé vers le cœur de la fleur y compris par la courbure incitative du tépale externe.
-

- Visite d’un bourdon sur une fleur d’iris de Hollande en 4 étapes : l’entrée en se glissant sous le style rabattu

-

- Le bourdon s’aplatit pour avancer dans le tunnel étroit ; il étire ses pattes arrière pour avancer

-

- Le bourdon déploie sa langue et va chercher le nectar au fond du tube au bout du tunnel

-

- Il ressort à reculons

L’abeille ou le bourdon va devoir forcer l’entrée entre le tépale externe et la branche pétaloïde du style rabattue ; elle est fermée ou rétrécie soit par le rapprochement très étroit des deux pièces ou soit par la présence de barbe ou d’une crête dressées qui poussent l’insecte vers le plafond. L’insecte cherche le nectar qui se trouve tout au fond du tube au bout du tunnel : il rampe donc dans le tunnel comme on peut le voir de profil chez certains iris aux fleurs assez « aérées ». Au passage, son dos poilu frotte sur l’anthère de l’étamine plaquée dans le creux du style courbé et récupère ainsi du pollen. Arrivé au bout du tunnel, l’insecte insère sa longue langue dans l’orifice étroit à la base de la fourche des trois styles et aspire le nectar. Le prélèvement accompli, il sort à reculons et s’envole.
-

- Deux visiteurs pour la même fleur : une abeille solitaire au premier plan et un bourdon dans le tunnel au second plan

-

- Bourdon en train de sortir à reculons après sa brève visite (quelques secondes)

-

- Les bourdons déplient leur longue langue car même au fond du tunnel, ils doivent forcer pour atteindre le nectar dans le tube

Barrières anti-selfie
A l’entrée, si le visiteur a auparavant déjà butiné une autre fleur, il peut porter sur son dos du pollen ; en entrant ainsi en force, il frotte son dos sur la face externe de la languette au bout du style déployé, le stigmate réceptif, et assure ainsi le transfert du pollen. A la sortie, avec du nouveau pollen issu de la fleur même en plus, il va plutôt frotter sur l’autre face du stigmate, non réceptive, ce qui diminue les chances d’autofécondation (dépôt de son propre pollen sur son propre pistil).
Ces protections ne s’arrêtent pas là ; le stigmate n’atteint sa maturité (et sa capacité à recevoir des grains de pollen) que un ou deux jours après l’ouverture, alors que les anthères commencent à s’ouvrir avant même l’éclosion, dans le bouton floral ; on parle protandrie (mâle d’abord). A l’éclosion, les filets des étamines encore assez courts portent la longue anthère à distance du stigmate au bout du style la séparant dans l’espace de ce dernier (herkogamie) ; les filets s’allongent un peu en fin de floraison et rapprochent l’anthère du stigmate mais celle-ci est alors pratiquement vidée de son pollen. . Enfin, barrière ultime, un système d’auto-incompatibilité de type génétique empêche la germination du pollen sur le stigmate de la même fleur. On voit donc que la fleur d’iris dispose d’une batterie de dispositifs subtils sélectionnés en faveur d’une pollinisation croisée entre fleurs de pieds différents.
-

- La structure de base triple peut inciter les visiteurs à ne butiner qu’une des sous-unités et aller voir ailleurs

-

- L’anthère reste en dessous du stigmate réceptif au sommet du style pétaloïde

Il a été montré sur l’iris des serpents (voir la chronique) que les abeilles ne visitent souvent qu’une des trois sous-unités de la fleur triple avant de passer à une autre fleur. Comme chaque fleur est relativement éphémère (quelques jours : voir la chronique), les probabilités s’en trouvent encore plus fortement limitées.
Importance inégale
Des chercheurs (3) ont étudié le rôle respectif des deux types de tépales (externes et internes) dans le processus de pollinisation en les coupant ou en les raccourcissant et en évaluant l’impact statistique sur les différentes étapes de la pollinisation d’un iris à crête (voir ci-dessus) : l’approche et l’atterrissage des butineurs ; le nombre total de grains de pollen enlevés par visite (en fonction de l’étroitesse du tunnel) et la production de graines qui traduit indirectement le dépôt de grains de pollen sur le stigmate.
Quand ou coupe ou raccourcit les tépales externes, le nombre d’approches et d’atterrissages de visiteurs diminue ainsi que le nombre total de grains de pollen enlevés et la production de graines. Plus ils sont raccourcis, moins il y a de visites effectives. Par contre, quand il y a une visite malgré tout, l’insecte enlève et dépose autant de pollen que dans une fleur non modifiée. Donc, la baisse du succès reproductif tient avant tout au nombre de visites en baisse ce qui confirme l’importance des tépales externes comme pistes d’atterrissage et guides visuels.
Quand on coupe ou raccourcit les tépales internes, ceux dressés au dessus de la fleur, le nombre d’atterrissages diminue mais uniquement si les tépales sont entièrement coupés ; la production de graines ne change pas preuve que les dépôts de pollen se font normalement. Le rôle de ces tépales internes semble donc bien moins déterminant dans le succès reproductif.

Fleur d’iris faux-acore vue de dessus : noter la différence de taille nette les tépales internes (dressés entre les styles) et le sépales externes avec le motif coloré
Ceci expliquerait pourquoi ces tépales internes sont nettement plus courts et moins élaborés que les externes. On peut se demander pourquoi au cours de l’évolution, ils n’ont pas encore plus régressé compte tenu du coût énergétique à produire de telles pièces. Les chercheurs émettent l’hypothèse d’un lien génétique entre les deux types de pétales au niveau de leur croissance : comme les externes subissent une forte pression en faveur d’une augmentation, la baisse des internes s’en trouverait limitée.

BIBLIOGRAPHIE
- Molecular Phylogeny of Crested Iris Based on Five Plastid Markers (Iridaceae). Jinyan Guo and Carol A. Wilson. Systematic Botany (2013), 38(4): pp. 987–995
- COMPARATIVE MICROMORPHOLOGY AND ANATOMY OF CRESTED SEPALS IN IRIS (IRIDACEAE). Jinyan Guo Int. J. Plant Sci. 176(7):000–000. 2015
- Functional differentiation in pollination processes between the outer and inner perianths in Iris gracilipes (Iridaceae). 2011. Canadian Journal of Botany: 17 84(1):164-171
A retrouver dans nos ouvrages
Page(s) : 406 à 411

Page(s) : 202 ; 440

