Arums et séparation des sexes
Arum italicum
Les arums au sens populaire partagent la même structure d’inflorescence reconnaissable au premier coup d’œil : un grand cornet membraneux coloré, la spathe (voir la chronique sur les bractées) et une sorte de massue charnue qui émerge au centre. Mais, pour le botaniste, ce nom vernaculaire d’arum recouvre en fait des dizaines de genres différents comme les grands arums blancs (genre Zantedeschia) ou l’arum serpentaire (genre Dracunculus : voir les chroniques sur ce genre) ; au sens strict, les « vrais » arums (les gouets en langage populaire) sont les espèces rattachées au genre scientifique Arum qui compte 26 espèces. Sur les quatre représentées en France, deux sont répandues dans une bonne partie du pays : l’arum maculé (Arum maculatum) et l’arum d’Italie (Arum italicum) (voir la chronique sur les gouets pour les critères distinctifs). Tous deux fleurissent en milieu de printemps et donnent l’occasion de découvrir un mode de reproduction très original et spectaculaire.
Remarque : Nous allons ici nous appuyer essentiellement sur l’arum d’Italie, aux « fleurs » plus grandes (et surtout pour lequel nous avons le plus de photos !) : les mensurations fournies concernent cette espèce (1).

Pied d’arum d’Italie fleuri en mai (Vendée)
Vraie fausse fleur
-

- Certaines spathes sont tachetées : sont-elles plus attractives ?

-

- Quand l’inflorescence émerge, la spathe est complètement enroulée sur elle-même

-

- Chez l’arum d’Italie, il y a souvent plusieurs inflorescences par pied

Pour les profanes, le cornet des arums est une fleur énorme et la massue qui émerge au centre fait bien penser au stigmate d’un pistil. Pour lever la supercherie, déchirons la spathe, cette bractée repliée sur elle-même en cornet, et entrons dans l’intimité de la chambre florale comme l’appellent les botanistes. Là, on découvre que la massue terminale, longue de 3 à 14cm est portée sur une tige courte (un pédoncule) de 1 à 4cm de long. Sur ce pédoncule, se succèdent quatre étages d’éléments qu’on a bien du mal à identifier au premier abord. Partons de la base pour remonter vers le spadice.
-

- Le spadice en forme de massue émerge de la spathe

-

- Il faut déchirer la spathe en cornet pour découvrir l’inflorescence étagée

-

- De haut en bas : fleurs stériles avec filaments ; fleurs mâles ; fleur stériles ; fleurs femelles

Un premier anneau (0,2 à 3,3cm de hauteur) assez marqué se compose d’éléments en forme de petites bouteilles ventrues surmontées d’un disque avec des papilles allongées ; chacune de ces boules contient des ovules : il s’agit donc de fleurs femelles et le disque papilleux fait office de stigmate chargé de capter le pollen. Ce sont elles qui, si elles sont fécondées, donneront des fruits charnus, des baies (voir la chronique communale sur les gouets).
Puis vient un anneau de 1 à 3mm de hauteur fait de une à deux rangées de filaments jaune crème, interprétés comme étant des fleurs femelles stériles transformées (pistillodes). Juste au-dessus, se trouve un anneau court (1cm au plus, composé de fleurs mâles très serrées, réduites à des étamines dont les anthères s’ouvrent par une fente en long pour libérer le pollen à maturité. Enfin, un dernier anneau de 2 à 5 rangées de filaments longs et raides, un peu sinueux et jaune crème. On les assimile à des fleurs mâles stériles transformées ou staminodes ; ils se trouvent juste au niveau de l’ouverture de la spathe, là où les deux bords du cornet se chevauchent : elles gardent en quelque sorte l’entrée en interposant une grille hérissée.
-

- Etage de fleurs mâles (productrices de pollen) encadré par deux étages de fleurs stériles filamenteuses
-

- Etage basal des fleurs femelles réduites à un ovaire
Disons que chez l’arum, il y a deux lits séparés dans la même chambre avec, en plus, une séparation de filaments entre les deux lits !
Fleurs de cristal
Le suivi du développement de cette inflorescence (1) a révélé, lors des premiers stades, un curieux phénomène : les fleurs mâles, les filaments situés au-dessus et le spadice (la massue) se recouvrent de masses globuleuses faites de cristaux d’oxalate de calcium. Ce dépôt de surface se produit avant que la spadice n’atteigne la maturité : l’inflorescence totale mesure alors moins de 10% de sa taille finale. Ces cristaux sont exsudés à la surface de l’épiderme de ces organes sans qu’il n’y ait aucune ouverture de cellules : leur dépôt sur les fleurs mâles n’a donc rien à voir avec l’ouverture tardive des anthères qui libère le pollen.
On sait que toute la plante contient de fortes quantités d’oxalate (voir la chronique sur les gouets) et ce dépôt externe serait un moyen d’éviter l’overdose interne qui finirait par devenir dommageable aux tissus ; secondairement, ces amas cristallins pourraient jouer un rôle répulsif vis-à-vis des herbivores car croquer ces masses toxiques très irritantes s’avère très ardu ! Peut-être aussi que cette accumulation précoce aide à la pollinisation mais on ne sait aucunement de quelle manière. Il s’agit peut-être tout simplement d’une forme d’excrétion !
Tromperie
Devant une structure aussi complexe, on ne peut que poser la question fatale : à quoi tout cela peut-il bien servir ? De toute évidence, il faut chercher du côté de la pollinisation, i.e. de la reproduction par transfert du pollen vers les stigmates des fleurs femelles pour assurer leur fécondation et leur transformation en fruits porteurs de graines. Cette inflorescence en cornet fermé fonctionne sur le mode du piège à pollinisateurs : elle attire certains insectes qui vont se retrouver piégés temporairement dans la chambre florale du cornet (plusieurs heures à plusieurs jours) et qui, en plus, ne recevront rien en retour puisque les fleurs femelles ne produisent aucun nectar nourricier ! C’est ce qu’on appelle la pollinisation par duperie (deception en anglais). Dans le cas des arums (et de nombreuses autres aracées : voir les chroniques sur la serpentaire), la plante attire des mouches et moucherons en imitant l’odeur et l’aspect des sites de reproduction de ces insectes qui pondent sur des matières en décomposition où leurs larves se nourriront. On parle de sapromyophilie (sapro = décomposition ; myo pour mouches et philie pour aimer).

Mouches piégées dans la chambre florale et attirées par l’odeur nauséabonde.
Chaud bouillant !
La floraison de cette inflorescence est accompagnée d’un processus bien connu chez les Aracées, le dégagement de chaleur ou thermogénèse ; on sait qu’elle provient d’un accroissement de la respiration cellulaire (par les mitochondries) qui normalement produit de l’énergie mais dans ce cas est détournée vers une émission de chaleur (voir aussi les chroniques sur la serpentaire). Dans le cas de l’arum d’Italie (2), on a mis en évidence quatre pics de chaleur pendant les deux à trois jours que durent la floraison et la pollinisation.
Un jour avant l’ouverture de la spathe complètement repliée jusqu’en haut, les fleurs mâles libèrent une première « bouffée de chaleur » entre 9H et 14H qui fait monter la température dans la chambre de plus de 7°C au-dessus de la température ambiante. On observe en même temps un léger mouvement des bords de la spathe de quelques millimètres : or, chez d’autres plantes, on a montré qu’un accroissement de la température favorisait les mouvements des feuilles ; les deux seraient donc liés ?
-

- Le dégagement de chaleur est tel qu’une condensation se forme sur le spadice au pic de chaleur
Le lendemain, la spathe s’entrouvre tandis que les fleurs mâles émettent un second pic de chaleur plus important (11°C en plus). Puis dans l’après-midi, un troisième pic de chaleur majeur apparaît, un vrai coup de chaud (+ 19°C qui affecte cette fois le spadice, la massue terminale qui se trouve donc hors de la spathe désormais ouverte. Il libère alors des composés volatiles qui diffusent dans l’environnement immédiat : une grosse dizaine de composés ont été identifiés dont de l’éthanol, du diméthyloctadiène et du méthyl butyrate. Le cocktail dégage une odeur… d’urine et de matières en décomposition qui va attirer les insectes pollinisateurs (voir ci-dessous). Ce coup de chaud n’affecte pas la chambre florale qui reste à une température légèrement au-dessus de la température extérieure et il y règne une ambiance olfactive plutôt sucrée. Le lendemain, alors que la pollinisation a eu lieu, un dernier pic de chaleur limité concerne à nouveau les fleurs mâles.
Çà sent mauvais !
Le suivi détaillé de pieds fleuris d’arums d’Italie dans la région de Toulouse sur deux sites distants de vingt kilomètres (2) a permis de déterminer et de quantifier les visiteurs attirés par ce piège odorant. Sur 174 spathes suivies, seulement 3 n’ont pas reçu de visites ce qui montre l’attractivité de ce dispositif. Chaque spathe contenait en moyenne de 3 à 7 insectes selon le site.
A 87% ce sont des Diptères, des mouches ou moucherons (termes très vagues qui recouvrent en fait des dizaines de familles différentes) avec largement en tête des Pyschodidés, des petits moucherons (1 à 5mm) très velus, ressemblant à de minuscules papillons de nuit ; deux espèces prédominent mais leur importance relative varie d’un site à l’autre. Les espèces du genre Psychoda se montrent les plus efficaces dans le transport du pollen sur leurs pattes et leur tête. Leurs larves vivent dans les eaux usées chargées en matière organique en décomposition. On a aussi capturé des chironomes (qui ressemblent à des moustiques), des sciaridés (les « mouches des terreaux ») et des sphaerocéridés (petites mouches dont les larves vivent dans les excréments, l’humus ou les végétaux pourrissants) ; l’arum d’Italie se comporte donc en généraliste mais à l’intérieur d’un cercle bien délimité d’espèces saprophages. De toute évidence, c’est bien l’odeur dégagée par le spadice qui leurre ces insectes qui croient trouver à l’intérieur de cette chambre fermée un site de ponte avec des matières en décomposition.
Piège doré
Ces insectes sont donc attirés au cours de la première journée de floraison au moment du coup de chauffe du spadice ; ils entrent par l’ouverture du cornet en se faufilant entre les filaments du cercle supérieur. A ce moment-là, seules les fleurs femelles sont mûres et réceptives ; ainsi, si ces moucherons ont auparavant visité une autre inflorescence d’arum (dans laquelle ils ont dû être piégés déjà !), ils transportent sur eux du pollen extérieur et vont le déposer sur les stigmates des fleurs femelles en circulant dans la chambre florale à l’ambiance chaude et sucrée, donc favorable à leur séjour mais sans leur apporter ni nourriture, ni site de ponte ! S’ils cherchent à fuir, ils le font en volant et se heurtent alors à la barrière des filaments raides et hérissés qui les piège.
-

- De loin, on a l’impression que même si la spathe est repliée il y a un passage

-
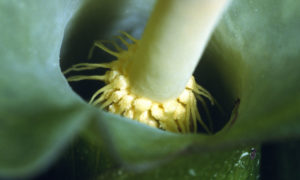
- De près, par dessus, on voit que les filaments forment une grille qui empêchent le départ en vol des moucherons qui sont entrés
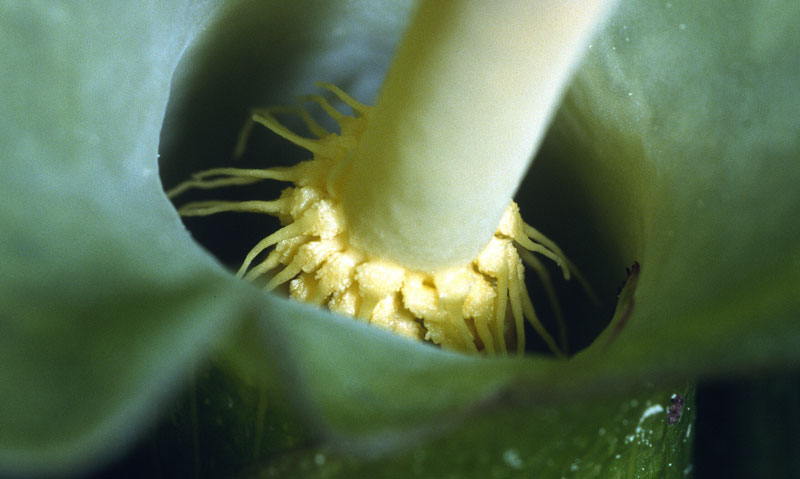
Ils vont ainsi rester plusieurs heures, voire plus d’une journée complète dans cette chambre, le temps que les fleurs mâles mûrissent et commencent à libérer leur pollen tandis que les fleurs femelles ne sont plus réceptives. Ainsi, ils peuvent prendre en charge du nouveau pollen. Le jour suivant, les filaments supérieurs flétrissent libérant le passage vers la liberté … Juste le temps de se faire alpaguer par une autre inflorescence d’arum qui entre en maturité !
-

- Du pollen a été déposé par des mouches (on les devine en arrière-plan) sur les fleurs femelles

A la séparation spatiale des fleurs mâles et femelles s’ajoute donc une séparation temporelle avec priorité au sexe femelle d’abord : on parle de protogynie, un dispositif qui limite considérablement les possibilités d’autofécondation (entre fleurs de la même inflorescence).
Efficace ?

Certaines années, si la météo est favorable, la reproduction peut être très réussie (Puy St Jean ; Artonne 63)
Chaque épi floral d’arum d’Italie compte en moyenne une soixantaine de fleurs femelles qui contiennent chacune environ cinq ovules ; potentiellement, une inflorescence pourrait donc produire plus de 300 graines si tous les ovules étaient fécondés. En réalité, seulement 66% des fleurs femelles se transforment suite à la pollinisation ; mais ensuite, nombre d’infrutescences avortent en cours de route : plus de 80% d’entre elles, ce qui est classique chez les Aracées ! Aucune inflorescence ne développe tous ses ovules même si elle a piégé des moucherons ad hoc : la pollinisation ne semble donc pas être le facteur limitant. La plante doit produire ses fruits en piochant dans les réserves du rhizome (voir la chronique sur les gouets) déjà bien entamées par la poussée des feuilles et la production des fleurs. Il est possible que s’effectue un avortement sélectif permettant le développement de moins de graines mais de meilleure qualité et donc plus viables.
-

- Epis de fruits en cours de maturation

-

- Epis de fruits mûrs très voyants

-

- Sur certains épis, on voit qu’un certain nombre de fruits sont très réduits et mal formés.

D’autre part, compte tenu du mode de pollinisation, on aura compris que la non synchronisation de la floraison entre pieds différents est importante pour que le système marche : un moucheron qui sort d’une inflorescence vieille de deux jours doit rencontrer une inflorescence juste épanouie pour que la pollinisation se fasse ; quand des périodes de temps froid et humides sévissent, cela bloque le processus et ensuite toutes les inflorescences mûrissent en même temps ce qui serait préjudiciable au succès de la pollinisation !
Des expériences d’ensachage d’inflorescences avec de la gaze montrent que l’autofécondation fonctionne très peu ce qui est logique vu la séparation spatiale des deux types de fleurs ; même si les fleurs mâles se trouvent au-dessus des fleurs femelles, le pollen qui tombe se trouve intercepté par le cercle inférieur de filaments.
BIBLIOGRAPHIE
- Development of the flower and inflorescence of Arum italicum (Araceae). Denis Barabé, Christian Lacroix, and Marc Gibernau. Can. J. Bot. 81: 622–632 (2003)
- Pollination ecology of Arum italicum (Araceae). J.ALBRE, A. QUILICHINI ; M. GIBERNAU. Botanical Journal of the Linnean Society, 2003, 141, 205–214
A retrouver dans nos ouvrages
Page(s) : 128-129

Page(s) : 46-47

