Classer les interactions entre espèces : une chimère ?
Aucune espèce ne vit seule dans son environnement : elle interagit avec celui-ci mais aussi avec les autres espèces présentes qui forment avec elles une communauté. On étudie ces interactions entre paires d’espèces. Les interactions interspécifiques sont une des clés de fonctionnement et de structuration des écosystèmes et aussi un des moteurs de l’évolution via les pressions sélectives dont elles font l’objet. Depuis longtemps, en écologie, on a cherché à catégoriser, à classer, ces interactions entre espèces soit en prenant en compte leur mode de fonctionnement, soit en s’appuyant sur leur intensité, leur durée et leur orientation. Nous allons voir historiquement les différentes classifications proposées et leurs limites pour terminer par une question paradoxale : les interactions sont-elles vraiment classables en catégories finies ? Cette chronique doit beaucoup à l’introduction de J.L Bronstein dans son ouvrage sur le mutualisme paru récemment (1).
La grille des interactions
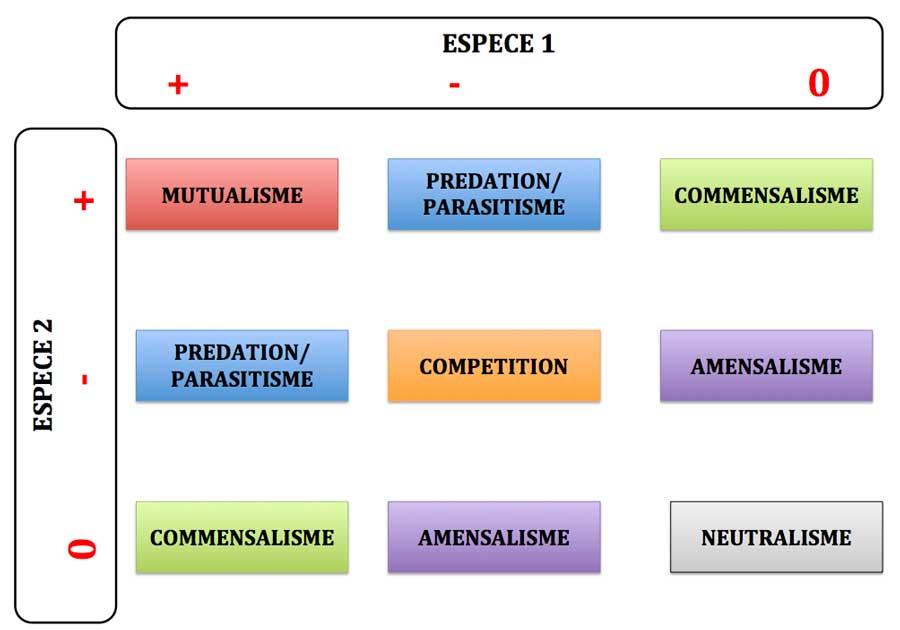
Dans la plupart des ouvrages ou sites de vulgarisation, on distingue six grandes catégories d’interactions selon leurs effets nets sur chacun des deux participants ; pour cela, on distingue trois grands types d’effets entre espèces :
– « + » : bénéfique pour le partenaire considéré
– « – »: nocive, négative pour le partenaire
– «0 » : neutre, sans effets.
La présentation classique est celle d’un tableau à double entrée avec les deux espèces (1 et 2) en interaction et les trois états du critère effet (+, – , 0) : dans chaque case du tableau, on peut ainsi définir un couple d’effets en interaction qui permet de définir les six grandes catégories (avec une symétrie par rapport à la diagonale).
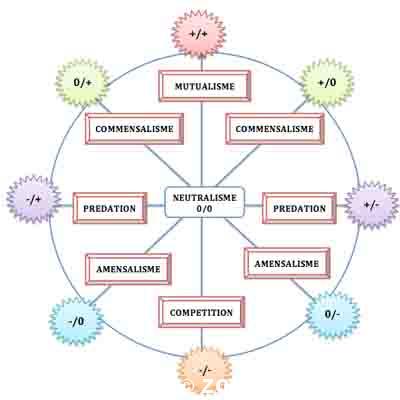
C’est ce qu’on appelle la grille des interactions, conçue historiquement en 1947 par E. Haskell (1906-1986), un scientifique atypique qui a dédié sa vie à tenter d’unifier toutes les connaissances humaines en une seule discipline ! Sa grille s’appliquait en fait à un système de classification des comportements sociaux humains. Cette genèse est particulièrement intéressante d’un point de vue épistémologique car les interactions écologiques ont toujours cristallisé sur elles ce parallèle ce qui a souvent brouillé leur analyse et leur perception. Cette grille fut donc reprise en 1953 par E. Odum, un des pères de l’écologie des écosystèmes, dans son ouvrage de référence Fundamentals of Ecology et, depuis, elle fait autorité tout en ayant subi diverses tentatives de remaniement.
Les six grandes
Donc, en lisant cette grille, on peut distinguer six grandes catégories d’interactions, symbolisées chacune par une paire d’effets sur les deux espèces partenaires :
– le mutualisme ou interaction +/+ : les deux espèces tirent un bénéfice de leur interaction
– la prédation et le parasitisme ou interaction +/- : une espèce tire profit tandis que l’autre est soit tuée (prédation) ou nettement affectée (parasitisme)
– le commensalisme ou interaction +/0 : l’interaction est bénéfique pour une des deux espèces tandis que l’autre n’en tire aucun avantage mais aucun inconvénient non plus
– la compétition ou interaction -/- : les deux espèces souffrent de leur interaction réciproque
– l’amensalisme ou interaction -/0 : une espèce agit de manière négative sur une autre mais sans pour autant en tirer aucun profit pour elle-même
– le neutralisme ou interaction 0/0 : les deux espèces interagissent mais sans aucun effet l’une sur l’autre.
Cette dernière catégorie est souvent omise ou non développée pour la simple raison, qu’en pratique, elle est quasiment indémontrable et il n’est pas sûr qu’elle existe vraiment dans la réalité ! En fait, c’est une interaction dans laquelle il n’y a pas d’interaction !!! Par contre, elle sert de de point de repère pour graduer les autres qui s’en rapprochent plus ou moins.
On voit se dessiner deux grands ensembles :
– les interactions mutualistes (ou positives) bénéfiques pour au moins un des deux partenaires : mutualisme et commensalisme
– les interactions antagonistes négatives pour au moins un des deux associés : prédation/parasitisme, amensalisme, compétition
Evidemment, à l’intérieur de chacune de ces catégories, on est amené à distinguer diverses sous-catégories ou variantes qui seront développées dans les chroniques consacrées à chacune de ces six catégories ; c’est pour cela qu’on parle par exemple des mutualismes ou des commensalismes pour signifier cette diversité.
NB : Par exemple, pour approfondir le commensalisme, aller dans l’alphabet du site à la lettre C et voir l’en-tête commensalisme qui regroupe plusieurs chroniques.
Une grille carcan
Dès les années 1980, cette grille fit l’objet de fortes critiques inhérentes à sa « structure ». D’abord, que signifient exactement les effets plus, moins ou zéro ? L’effet (+) plus veut-il dire que la population d’une espèce augmente quand elle est en interaction avec l’autre ou que la survie et le succès reproductif de ses membres s’améliore plus en présence de l’autre ? A partir de quelle degré d’augmentation ou de baisse considère t’on que l’effet est réellement positif ou négatif ? Certains des termes utilisés comme prédation, parasitisme ou compétition font appel à des mécanismes alors que le classement s’appuie sur les effets. Enfin, et c’est là sans doute le principal écueil de cette grille, c’est de faire des interactions quelque chose de rigide, statique, bien défini. Or, les faits indiquent tout le contraire : une même interaction peut avoir des effets différents selon le contexte (l’environnement dans lequel elle a lieu), les densités respectives des deux espèces et évoluent dans le temps (comme par exemple dans les successions végétales). A long terme, sous la pression sélective, les interactions se transforment et peuvent même changer de nature. Des cases contraintes et bien délimitées ne peuvent répondre à toutes ces variations et nuances. Il n’empêche que cette grille continue à être utilisée pour sa commodité mais il ne faut pas oublier, si l’on s’en sert, de la nuancer et de la critiquer très fortement !
La boussole des interactions
Il fallait trouver une représentation qui prenne en compte en fait le continuum existant entre toutes ces interactions avec tous les intermédiaires et évolutions possibles. Bizarrement, c’est le même E. Haskell qui en 1949 suggéra une seconde version de sa grille sous la forme d’un cercle et qu’il surnomma « la boussole des coactions ».
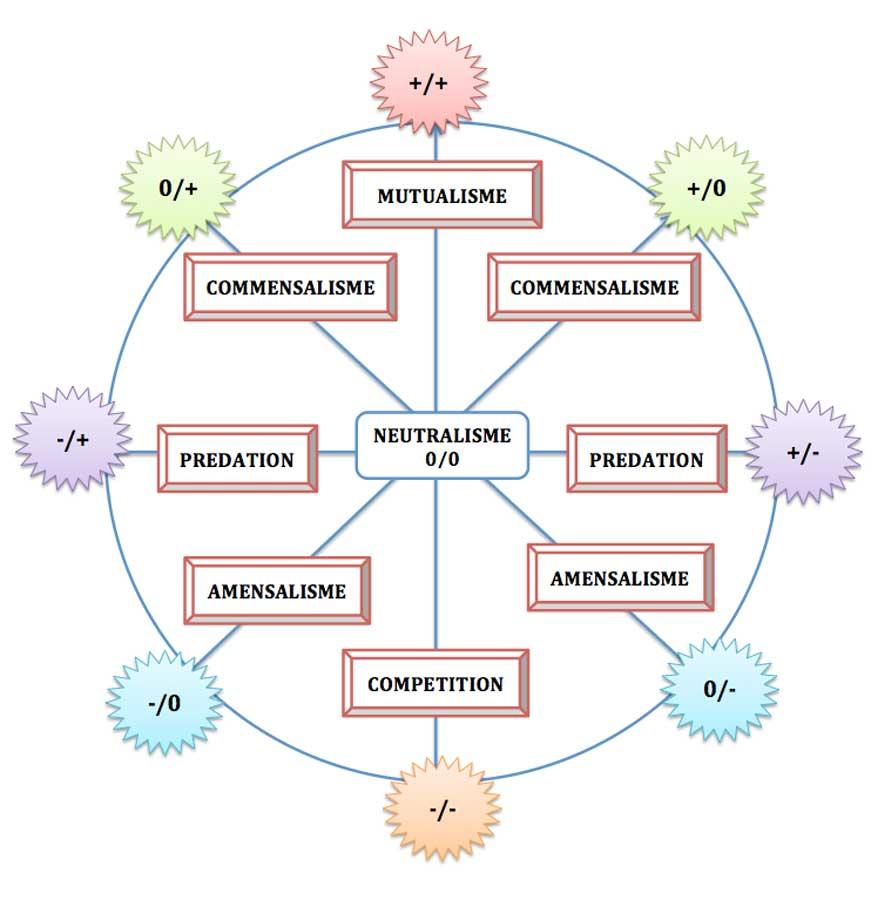
Cette « boussole » présente un avantage majeur par rapport à la grille : celui de la flexibilité et de la gradation. La circonférence représente « la surface nulle » où l’effet de l’interaction sur le système écologique représenté par les co-acteurs est nulle ; si on se déplace vers l’intérieur du cercle, il y a des pertes et si on sort du cercle, il y a des gains de production du système. En se se déplaçant le long du cercle, on peut passer graduellement d’une catégorie à une autre ; or, c’est bien ce qui se passe souvent (voir par exemple la chronique sur la diversité des commensalismes où de nombreux cas intermédiaires commensalisme/mutualisme et commensalisme/prédation (par ex. les pinnothères dans les moules) sont détaillés).
Interactions instables

Un exemple de chenilles défoliatrices (Yponomeuta sp) sur une colonie de Prunelliers au printemps : elles recouvrent les arbustes de leurs toiles communautaires et dévorent toutes les feuilles
Un exemple précis va illustrer à la fois la flexibilité de cet outil et celle des interactions elles-mêmes d’une année à l’autre (2). Cette étude américaine porte sur l’interaction très classique (herbivorie, autrement dit une variante de la prédation) entre un insecte phytophage, un papillon de nuit, la livrée des forêts (Malacosoma disstria) dont les chenilles font des nids en forme de tente sur les peupliers trembles américains (Populus tremuloides) ; elles consomment les feuilles : ce sont des chenilles défoliatrices. Certaines années où leurs populations augmentent considérablement, les peupliers peuvent être entièrement défoliés mais ils refont une seconde poussée de feuilles en fin de printemps. Contrairement à une idée reçue, l’effet de ces chenilles sur les arbres n’est pas toujours négatif ; si la défoliation ne touche que 5 à 30% du feuillage, la production primaire de l’arbre n’est pas affectée et, fait le plus surprenant, cela peut même accélérer sa croissance ! Il semble que des massifs de tremble défoliés 2 ou 3 ans de suite présentent en fait une meilleure croissance en hauteur et une légère augmentation de la croissance en diamètre du tronc. On explique cet effet paradoxal ainsi : la circulation des nutriments minéraux essentiels tels que nitrate, phosphore et potassium serait stimulée ou la répartition de la lumière et de l’humidité dans le feuillage serait plus équitable. Les défoliations entraînent un enrichissement de la litière en ces minéraux de 20 à 200% à cause des cadavres des chenilles, de leurs mues et de leurs innombrables excréments qui tombent au sol ainsi que les débris de feuilles échappés ! On a affaire ici à une relation trophique entre un « petit » consommateur et un énorme producteur primaire : cette différence de niveaux explique aussi une part de ces décalages observés.
-

- Toiles de Yponomeuta sur des prunelliers

-

- Les chenilles sont présentes par milliers et se protègent grâce à leur toile commune

-

- En fin de printemps, les chenilles se sont nymphosées ; contre toute attente, les prunelliers repartent aussitôt et refont une nouvelle génération de feuilles et arrivent même à se reproduire.

-

- Chenilles de Malacosoma neustria, une espèce européenne proche de celles étudiée aux U.S.A.

En associant un modèle de simulation et des mesures de production primaire sur le terrain, les deux chercheurs ont pu ainsi reconstituer sur plus d’une dizaine d’années l’évolution de cette interaction chenille/tremble en termes d’impact sur la production primaire de l’arbre ; ainsi, ils ont pu année après année qualifier le type d’interaction qui peut varier de commensalisme (les chenilles mangent les feuilles mais l’effet est nul sur les feuilles) à prédation-parasitisme alimentaire (pullulation massive de chenilles qui diminue la production) à des situations de quasi-mutualisme (amélioration de la croissance) ! On peut avec ces valeurs positionner précisément pour chaque année l’interaction sur la boussole et matérialiser sa trajectoire dans le temps !
Bénéfices et coûts
La morale de cet exemple (et il est loin d’être le seul), c’est que pour toutes les interactions, quelles qu’elles soient, il faut prendre en compte autant les bénéfices que les coûts pour les deux espèces impliquées que l’effet soit positif ou négatif. Dans l’exemple précédent, la défoliation a un coût certain pour l’arbre mais aussi une part de bénéfice. De la même manière, dans les relations mutualistes on dit que les effets sont positifs des deux côtés ; pour autant, cela ne signifie pas qu’il n’y aucun coût pour chacun des deux partenaires mais que les bénéfices l’emportent sur les coûts. Ainsi, les fleurs qui par leurs couleurs attirent des pollinisateurs risquent aussi d’attirer des insectes herbivores (une fleur çà se mange aussi !) et de voir la consommation de leurs précieux organes reproducteurs (coûteux à produire) augmenter. Ces équilibres bénéfices/coûts évoluent sans cesse dans le temps et d’un lieu à l’autre selon le schéma de la mosaïque géographique sous l’effet des pressions sélectives.
Rien n’est simple et encore moins simpliste : c’est cela qui est magique en science ; on croit avoir compris mais non, c’est bien plus compliqué que cela, presque hors de portée de nos cerveaux !
BIBLIOGRAPHIE
- MUTUALISM. Ed. by J. L. Bronstein. Oxford University Press. 2015
- Phytophagous insects as regulators of forest primary production. W. J. Mattson ; N. D. Addy. Science, New Series, Vol. 190, 1975 ; p. 515-522