Ancolies : des éperons qui aiguillonnent la biodiversité
Aquilegia

Eperons dressés chez une ancolie cultivée : la fleur aux cinq colombes.
Si on analyse l’arbre du vivant et ses innombrables rameaux, on constate que dans divers genres ou familles, on observe une très forte diversification avec de nombreuses espèces relativement différentes sur le plan morphologique ou écologique tout en étant issues d’un même ancêtre commun ; on parle de radiation évolutive comme si une explosion de biodiversité s’était produite au sein de certaines lignées. Pour expliquer pourquoi certaines lignées seraient concernées et pas d’autres très proches, on fait souvent appel au concept d’innovation évolutive clé, un trait ou caractère nouveau apparu à l’origine de la lignée et qui permet aux espèces descendantes d’exploiter leur milieu de manière différente. Chez les plantes à fleurs, un exemple bien connu et étudié depuis des décennies concerne les ancolies auxquelles nous avons déjà consacré une chronique : la fleur aux cinq colombes, titre évocateur de l’existence chez ces fleurs de cinq parties portant chacune un éperon. Y a t’il un lien entre la diversité des ancolies et la présence de ces organes originaux que sont les éperons : autrement dit, l’acquisition d’un éperon a t’elle été une innovation clé dans leur lignée ?
Proximité
-

- Ancolie des neiges du Bhutan

-

- Ancolie de Corée

Nombre d’indices pointent vers une évolution rapide et récente dans la lignée des ancolies (le genre Aquilegia) au sein de la famille des Renonculacées, ce que les scientifiques appellent une radiation évolutive, une évolution rapide dans une lignée issue d’un même ancêtre commun avec une forte diversification et disparité : de nombreuses espèces différant par leur écologie et/ou leur morphologie.
-

- L’ancolie du Colorado (A. caerulea) (ici, un cultivar) possède plusieurs formes colorées selon les populations

-

- Fleur de face : le port horizontal et les longs éperons fins indiquent une fleur visitée par des sphinx

On dénombre une bonne soixantaine d’espèces d’ancolies toutes dans l’Hémisphère Nord de l’Eurasie à l’Amérique du nord et on observe un fort degré d’inter-fertilité entre ces espèces avec des hybridations fréquentes dans les zones de sympathie (quand deux espèces se côtoient) ; cette caractéristique a d’ailleurs été largement exploitée par les horticulteurs pour créer une foule d’hybrides et de nouvelles variétés : ceci indique qu’on a affaire à des espèces génétiquement relativement semblables, ayant donc évolué rapidement et sans doute récemment. Ces niveaux de divergence des séquences génétiques très bas se retrouvent aussi bien dans l’ADN du noyau (ADN ribosomal) que dans celui des chloroplastes, preuve supplémentaire d’une évolution rapide et récente.
-

- Une autre ancolie américaine : l’ancolie dorée (ici un cultivar) aux très longs éperons fins

-

- De face, on note sur le pétale du haut l’entrée vers l’éperon ; il s’agit là encore d’une ancolie pollinisée par des sphinx

Disparité
Si les ancolies se reconnaissent facilement au premier coup d’œil de part la structure très particulière de leurs fleurs à cinq éperons, elles n’en diffèrent pas moins fortement entre espèces par une foule de caractères morphologiques au niveau de ces mêmes fleurs (et très peu au niveau des caractères végétatifs). Les différences portent sur les couleurs des fleurs avec des dominantes soit bleues, soit jaunes soit rouges ou blanches, sur la forme des éperons (longs, courts, droits, courbés, crochus, …), l’orientation des fleurs (dressées, horizontales ou pendantes) et sont associées étroitement aux types de pollinisateurs dominants propres à chaque espèce : bourdons, abeilles, papillons nocturnes dont les grands sphinx ou colibris en Amérique du nord.
-
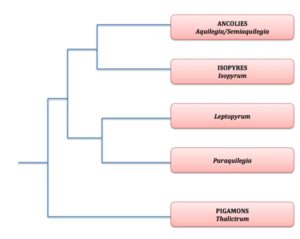
- Arbre de parentés des genres proches des ancolies

-

- Isopyre faux-pigamon : il ressemble à l’anémone sylvie et fréquente les sous-bois humides

-

- Pigamon glauque à fleurs jaunes : les fleurs sont pratiquement -réduites aux seules étamines !

-

- Pigamon à feuilles d’ancolie de nos massifs montagneux

Si on regarde ce qui se passe dans le genre le plus étroitement apparenté aux ancolies (groupe-frère), le genre Isopyre (Isopyrum) on constate que ce dernier ne regroupe que quatre espèces bien qu’il soit réparti lui aussi sur une vaste aire de l’Hémisphère nord. Dans ce genre, les fleurs sont rayonnantes et régulières et la pollinisation est assurée par une foule d’insectes généralistes. On voit donc se dessiner un scénario évolutif où l’apparition d’éperons dans une lignée a permis la spécialisation envers différents pollinisateurs et donc l’apparition de nombreuses espèces (spéciation) différant surtout au niveau de leurs caractères floraux. Ainsi, on peut poser sur ces éperons l’hypothèse qu’il s’agirait d’une innovation évolutive clé.

Pigamon de Delavay : les pigamons sont très proches des ancolies et possèdent pourtant des fleurs complètement différentes sans éperon.
Un dernier indice de forte diversification concerne l’écologie des ancolies que l’on peut rencontrer dans des milieux très divers allant des alpages des hauts sommets montagneux, aux forêts variées jusqu’aux abords des sources en plein désert américain. Pour autant, vu l’étendue de l’aire de répartition, il ne s’agit pas d’une invasion de nouvelles niches écologiques mais bien d’une diversification morphologique en lien avec la pollinisation.
L’effet éperons
Ca change quoi pour une plante à fleurs d’avoir des éperons ? Allons voir ce qui se passe dans d’autres genres non apparentés et eux aussi dotés d’éperons pour mieux comprendre.Dans l’immense famille des orchidées, plusieurs genres possèdent un éperon dans leur fleur comme le genre Platanthère (Platanthera) représenté en France continentale par deux espèces très proches dont la platanthère à deux feuilles (P. bifolia) remarquable avec son long éperon très fin qui prolonge la fleur en arrière.
-

- Platanthères à deux feuilles dans une prairie

-

- Epi de platanthère à deux feuilles

-

- Fleur vue de face avec au centre le passage vers l’éperon, encadré par les deux masses de pollen qui se collent sur la tête du pollinisateur

Dans les années 1970, L.A. Nilsson, botaniste suédois, avait travaillé sur cette espèce en réduisant expérimentalement la longueur de son éperon ; il avait ainsi montré que les plantes avec des éperons plus courts produisaient moins de fruits que celles avec des éperons plus longs ; autrement dit, la longueur de l’éperon influe fortement sur le succès reproducteur de l’espèce. En Scandinavie, par exemple (1) elle est surtout visitée et pollinisée par des sphinx de plusieurs espèces dont le petit sphinx de la vigne ou le sphinx du liseron, attirés à la nuit tombée (papillons plutôt crépusculaires) par la couleur blanc pur des fleurs et le fort parfum vanillé qu’elles répandent à ce moment-là. Or, ces différentes espèces de sphinx possèdent une longue trompe dont la longueur peut varier de 18 à 45mm selon les espèces locales (1) ; pour que la visite d’un sphinx soit profitable à cette orchidée (pollinisation effective), il faut que la longueur de la trompe soit la plus proche possible de celle de l’éperon ce qui oblige le sphinx à appuyer sa tête à l’entrée de la fleur et ainsi à récolter les deux masses de pollen qui l’encadrent.
-

- Sphinx du liseron : une grande espèce

-

- Chrysalide de sphinx du liseron : la trompe est tellement longue qu’elle dépasse de la chrysalide !

-

- Petit sphinx de la vigne

-

- Grand sphinx de la vigne, autre espèce proche

Or, la répartition de ces diverses espèces de sphinx varie selon les milieux : on constate que les platanthères des bois avec un long éperon et celles des prairies avec un éperon plus court en moyenne ne sont pas visitées par les mêmes espèces et que la longueur de l’éperon est corrélée à celle de la longueur de la trompe des sphinx visiteurs.

Le long éperon de la platanthère à deux feuilles : le nectar s’accumule tout au bout
D’autres exemples connus vont dans le même sens : la morphologie des éperons est intimement associée au type de pollinisateur.
Isolement

Les longs éperons très élégants de l’ancolie dorée avec leur extrémité pleine de nectar qui n’est accessible qu’à des visiteurs avec une très longue trompe !
Les éperons apportent donc en fonction de leurs variantes morphologiques la possibilité de spécialisation envers tel ou tel type de pollinisateur ce qui constitue un facteur d’isolement reproducteur, i.e. que les échanges de gènes ne se font plus qu’entre plantes partageant les mêmes pollinisateurs précis. Or, un exemple précis de ce processus a été longuement étudié à propos de deux espèces d’ancolies des montagnes de l’Ouest de l’Amérique du Nord depuis les années 1950 : l’ancolie rouge (A. formosa) aux fleurs penchées d’un beau rouge cramoisi, pollinisée surtout par des colibris et l’ancolie de Colville (A. pubescens) aux fleurs blanc crème horizontales visitées par des sphinx. Ces deux espèces se côtoient dans les hautes montagnes de Californie sur un long gradient d’altitude et réussissent pourtant à se maintenir comme espèces séparées avec un taux d’hybridation réduit. L’explication de la séparation de ces deux espèces sympatriques a fait l’objet d’un vif débat scientifique : certains avancent une explication écologique avec des types de sol différents associés à chaque espèce et qui les maintiendraient ainsi séparées dans l’espace ; d’autres mettent en avant les fameux éperons et apportent de sérieux arguments ! L’étude détaillée de la longueur et de l’orientation des éperons en fonction de l’altitude montre des transitions brutales : les éperons semblent subir une sélection contre les variations qui s’écartent du modèle type, celui de l’ancolie rouge associé aux colibris et celui de l’ancolie de Colville associée aux sphinx ; le comportement très spécifique de ces deux types de pollinisateurs doit jouer contre les variations alternatives de longueur et d’orientation. Il reste une hypothèse synthétique : les caractères floraux seraient étroitement liés aux caractères physiologiques (liens avec le sol) mais ceci semble très peu probable.
-

- Ancolie vulgaire : l’espèce indigène la plus répandue dans les milieux forestiers

-

- Fleur avec ses cinq éperons dressés

-

- Ancolie vulgaire cultivée visitée par un bourdon

-

- Ancolie vulgaire cultivée visitée par une abeille domestique. Fleurs pendantes et éperons courts et dressés sont typiques des ancolies à hyménoptères.

Des ancolies sans éperon
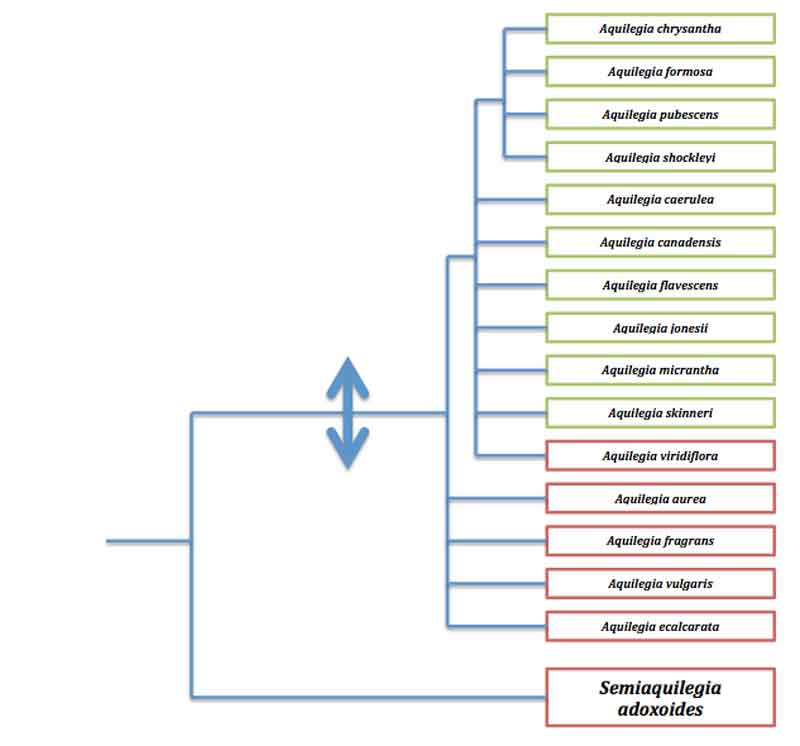
Arbre de parentés des ancolies ; en rouge : espèces eurasiatiques ; en vert : espèces nord-américaines. La double flèche indique l’apparition des éperons dans la lignée. D’après (2).
Globalement, tout semble converger vers l’hypothèse initiale des éperons comme innovation évolutive clé mais encore faut-il démontrer que les taux de diversification observés soient bien corrélés avec l’apparition de ces éperons. Or, au sein du groupe des ancolies, on connaît deux espèces originales … dépourvues d’éperon : l’ancolie sans éperon (A. ecalcarata) des montagnes de Chine et la semi-ancolie (Semiaquilegia adoxoides) des collines de Chine. Un arbre de parentés des ancolies incluant ces deux espèces atypiques apporte des éclairages intéressants. L’ancolie sans éperon se place auprès des ancolies avec éperon dont l’ancolie commune, l’espèce la plus répandue chez nous, ce qui indiquerait peut-être que chez cette espèce les éperons aient été perdus secondairement comme on l’observe régulièrement chez des mutants de formes cultivées ou ponctuellement sur des populations sauvages de certaines espèces. Par contre, la semi-ancolie occupe une position basale nettement divergente qui en fait une lignée ancestrale. Ceci permet d’avancer que l’éperon est apparu au début de la radiation des ancolies et qu’ensuite les espèces ont divergé très rapidement menant à la diversité actuelle ; cette coïncidence entre l’apparition de l’éperon et « l’explosion » des ancolies renforce donc l’hypothèse de l’innovation clé.
-
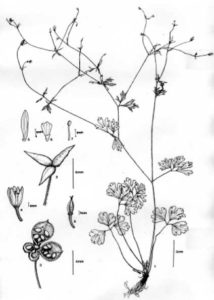
- Semi-ancolie (Flore de Corée)
-
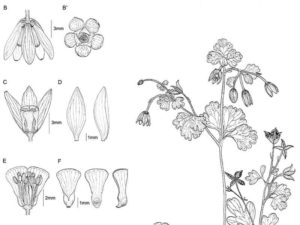
- Détails de la fleur de semi-ancolie, sans éperon.
L’étude du développement floral des ancolies (3) montre que la formation des éperons n’intervient que tardivement dans le développement de la fleur ; il se pourrait donc que un seul gène majeur contrôle la formation des éperons ; ceci expliquerait la facilité avec laquelle ces derniers ont pu varier et entraîner des changements de pollinisateurs et un certain isolement reproducteur favorisant l’émergence de nouvelles espèces.
Eperons boosters
Là encore, il est intéressant d’aller voir « ailleurs » dans d’autres groupes non apparentés avec des genres dotés d’éperon. On en connaît de nombreux exemples à travers l’arbre immense des plates à fleurs ; si on sélectionne les groupes dotés d’éperon et dont on connaît le groupe-frère (le plus proche parent) sans éperon, on constate que dans 7 cas sur 8, il y a plus d’espèces dans le groupe avec éperon ce qui conforte l’hypothèse de l’éperon « aiguillon de la diversification ».
-

- Fleurs de Pelargonium : le pédicelle rouge porte la fleur et renferme l’éperon « intégré et caché »

-

- On devine l’entrée de l’éperon en haut de la fleur ; noter aussi la tendance de la fleur à devenir irrégulière (zygomorphe) en lien avec l’éperon et la présence de taches colorées servant de guides à nectar qui balisent l’ouverture de l’éperon

-

- Géranium de Renard : la fleur n’a pas d’éperon (même caché) et est régulière

Il reste une exception notoire dans la famille des géraniacées, le genre bien connu Pelargonium (que nous appelons abusivement géranium) regroupe 280 espèces et leurs fleurs possèdent un long éperon ; par contre, le groupe-frère au sein de la famille (les genres Geranium, Erodium, Nonsonia et Sarcocaulon) compte près de 400 espèces alors que leurs fleurs n’ont pas d’éperon. Oui mais l’éperon des Pelargonium est très particulier : il est « invisible » ! En effet, il se trouve soudé au pédicelle de la fleur ce qui le cache au regard et l’observer demande une dissection florale ! Donc, toute modification de l’éperon impose une modification parallèle du pédicelle associé ce qui constituerait une contrainte forte sur l’évolution de l’éperon dans ce genre et aurait limité relativement sa diversification !

Fleur de Pelargonium en bouton : noter l’aspect épaissi du haut du pédicelle qui trahit la présence de l’éperon à l’intérieur
BIBLIOGRAPHIE
- Pollinator shifts and the evolution of spur length in the moth-pollinated orchid Platanthera bifolia. Elin Boberg et al. Annals of Botany 113: 267–275, 2014
- Rapid radiation due to a key innovation in columbines (Ranunculaceae : Aquilegia). S. A. Hodges. Chapitre 13 ; pp. 391-405 in Molecular Evolution and Adaptative Radiation. T.J. Givnish ; K. J. Systma. Cambridge University Press. 2000
- Floral nectar spurs and diversification. S.A. Hodges. Int. J. Plant. Science 158 (6 suppl.) : S81-S88 ; 1997
A retrouver dans nos ouvrages
Page(s) : pp. 580-82

Page(s) : pp. 580-81

Page(s) : pp. 88-89

