Sauges : des étamines basculantes, çà change tout !
Salvia

Inflorescence de sauge des prés 

Inflorescence opulente et ramifiée de la sauge sclarée
Les sauges, représentées par plus de mille espèces dans le monde, sont regroupées en un seul genre (Salvia) dans la famille des Labiées ou Lamiacées au sein de la tribu des Menthae qui compte 73 genres différents. Elles diffèrent fortement des 72 autres genres de cette tribu par un caractère très original : elles n’ont que deux étamines fonctionnelles au lieu de quatre classiquement et capables de basculer lors de la visite d’un pollinisateur. Ce dispositif unique avait été décrit dès la fin du 18ème siècle par C. K. Sprengel, un des premiers grands spécialistes de la pollinisation des fleurs avec son ouvrage fondateur « Le secret de la Nature découvert dans la forme et la fécondation des fleurs » (1793). Outre la beauté insolente et extravagante de nombre d’entre elles, les sauges continuent de fasciner les biologistes par la diversité des formes de leurs fleurs à deux lèvres, de leurs composés chimiques secondaires (dont des huiles essentielles) et des espèces animales assurant leur pollinisation. Depuis les années 2000, les sauges ont été retenues comme modèle d’approche interdisciplinaire à cause de cette structure florale hautement spécialisée et de leur vaste répartition sur plusieurs continents afin d’élucider les mécanismes d’apparition de nouvelles espèces (spéciation) dans ce genre. Pour rendre compte des résultats de ces travaux, nous allons nous appuyer sur deux espèces indigènes : la sauge des prés (S. pratensis), commune dans les milieux herbacées et la sauge sclarée (S. sclarea), méditerranéenne mais cultivée comme ornementale bien en dehors de sa zone originelle.

Sauges sclarées au jardin 
Pré sec à sauge des prés
Action-réaction

Pour comprendre le fonctionnement ce mystérieux levier, commençons par observer de l’extérieur ce qui se passe lors d’une visite d’insecte pollinisateur. Nous allons prendre comme exemple la sauge sclarée avec ses opulentes floraisons qui attirent comme visiteur principal les grosses abeilles charpentières noir bleutées ou xylocopes (voir la chronique sur ces insectes) ce qui facilite l’observation ! Néanmoins, il faut rester très attentif car la visite d’une fleur ne dure que … quelques secondes ! Le xylocope arrive face à la fleur et s’accroche lourdement sur la lèvre inférieure blanche plissée ; aussitôt, il enfonce sa tête dans le creux devant lui : c’est l’entrée rétrécie du tube floral de la corolle à la jonction avec la grande lèvre supérieure rose tendre. Ce qui intéresse le xylocope, c’est le nectar riche en sucrose produit tout au fond du tube de la corolle : il lui faut donc aller l’atteindre avec sa longue langue. Dès que la tête du xylocope bute sur l’entrée, on voit sortir de sous la lèvre supérieure en forme de casque deux étamines courbées. Elles basculent dans un mouvement gracieux et très rapide et viennent toucher le dos de l’insecte y déposant du pollen : les étamines basculantes, les voici ! Alors que le xylocope insiste en cherchant à enfoncer plus en avant sa tête, les anthères glissent sur son thorax vers l’arrière. Puis, dans un ultime « coup de rein » pour aller plus en avant, il redresse le bout de son abdomen en le cambrant fortement si bien que les anthères poursuivent leur frottement vers les premiers segments de l’abdomen. Le résultat se voit immédiatement sur le fond noir de l’insecte : le thorax et le haut de l’abdomen sont recouverts de pollen clair !

Position de départ : l’abdomen est rabattu 
Position finale : l’abdomen se cambre 
Noter l’abondant pollen sur le dos du xylocope
Finalement, la lèvre supérieure elle-même bascule un peu en arrière sous la pression ce qui fait sortir le long style courbé logé lui aussi sous celle-ci. Son stigmate terminal en forme de fourche vient toucher le xylocope sur le milieu de l’abdomen car le style porteur est plus long que les étamines : dans ce dernier temps, le stigmate peut donc récolter quelques grains de pollen sur le dos du xylocope s’il le touche. Il y a un subtil décalage dans la maturation du stigmate et des étamines : au début, le stigmate n’est pas réceptif alors que les anthères libèrent du pollen ; plus tard, le style s’allonge et met le stigmate plus en avant tandis que les étamines ne produisent presque plus de pollen. Ainsi, le stigmate aura plus de chance de toucher du pollen porté sur le dos du xylocope venant d’une autre fleur, ce qui facilite la pollinisation croisée.
Fleur bilabiée

On peut très facilement imiter le travail des xylocopes avec un crayon que l’on introduit de face dans la gorge de la corolle : on voit alors ce mouvement de bascule des étamines et du style, une belle expérience à faire avec des enfants ! Mais pour découvrir le mécanisme qui actionne ce mouvement il va falloir examiner l’intérieur de la fleur. Prenons cette fois comme exemple l’autre sauge, la sauge des prés aux fleurs plus faciles à disséquer et plus colorées pour la prise de vue. Détachons donc une fleur en coupant son court pédicelle qui la rattache à la tige, à l’aisselle d’une feuille bractée et en compagnie de quelques congénères réunies en étage (pseudo-verticilles). Le calice plissé, verdâtre, à dents épineuses, fortement nervé, couvert de poils glanduleux odorants, lui-même à deux lèvres, englobe la base de la corolle. Si on tire doucement sur celle-ci en la saisissant par la lèvre inférieure, elle se détache facilement d’un bloc. Il reste le long style courbé et son stigmate bleuté fourchu qui sort du calice. En déchirant ce dernier, on trouve l’ovaire en quatre parties séparées arrondies et le style : il émerge à la base (gynobasique) des quatre éléments qui vont donner les futurs fruits secs (tétrakènes). L’ovaire repose sur une masse charnue jaunâtre, un nectaire qui produit le nectar tant recherché des xylocopes.

Fleur de profil avec le calice vert 
Fleur au stade où le style s’est allongé 
Fleur de face telle que la voit un insecte qui arrive : entrée rétrécie et plate forme d’atterrissage 
Fleur disséquée révélant les deux étamines
La corolle détachée comprend un tube allongé terminé par les deux grandes lèvres opposées : la supérieure en forme de faux et l’inférieure, trilobée et plissée à sa base. De face, on voit que l’entrée du tube reste relativement resserrée par ces plis et on devine deux tiges claires qui ferment encore plus le passage : il faut va falloir maintenant déchirer à son tour délicatement la corolle. Ainsi, on découvre enfin les deux étamines complètement cachées jusque là et fixées en haut du tube, à la jonction avec la lèvre inférieure. On reconnaît facilement les anthères qui libèrent le pollen : chacune d’elles est portée par une longue tige arquée mais vers la base, tout se complique et on découvre une structure complexe avec des appendices : pas simple du tout et on s’y perd très vite !

Anthères à une loge ouverte avec le pollen et soutenues par le connectif 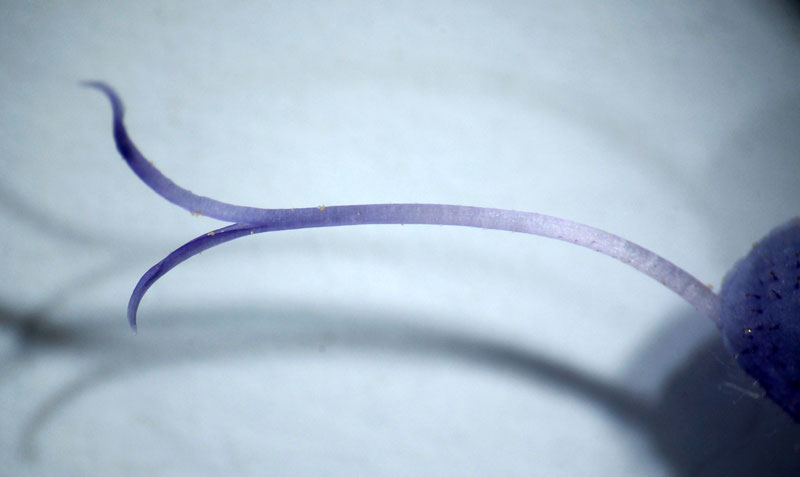
Style et stigmate bifide 
Style inséré à la base de l’ovaire vert 
+Ovaire en 4 éléments surmontant le gros nectaire jaune
Métamorphose

Schéma d’une étamine 
Filament vertical (fixé à la corolle) qui s’articule avec le grand connectif 
Les deux étamines détachées sans leurs filaments 
Sorte de « ligament » brun qui unit les deux cuillères à leur base
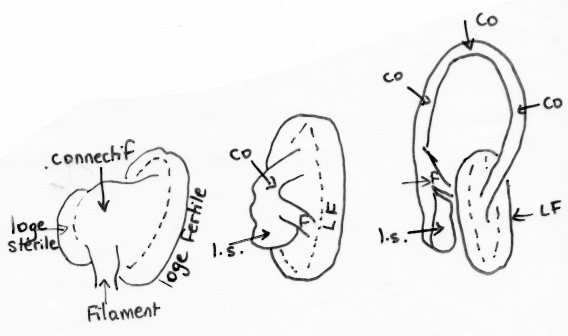
Nous entrons maintenant dans le dur du sujet : interpréter cette structure si particulière des deux étamines. On ne pourra comprendre la suite que si on a en tête la structure classique d’une étamine « normale » : une tige mince ou filet qui porte une anthère à deux loges ou thèques remplies de pollen et réunies entre elles et au filet par un très court connectif le plus souvent quasi invisible. Bien, maintenant passons aux étamines de la sauge : le longue tige courbée qui porte l’anthère n’est pas un filet mais le connectif qui réunit normalement les deux loges ; effectivement, l’anthère n’a qu’une loge fertile pleine de pollen et l’autre, stérile, s’est trouvée séparée par cette croissance anormale du connectif ; on la retrouve tout à la base sous la forme d’un appendice prolongeant une cuillère aplatie : méconnaissable ! Juste au-dessus de cette cuiller, vient s’articuler une tige droite, fixée sur le tube de la corolle : voici le vrai filet, très court par contre ! Si on dissèque délicatement la corolle, on découvre que ces deux étamines, en plus d’êtres fixées chacune par un filament, se trouvent en plus rattachées l’un à l’autre à la base des cuillères par une sorte de ligament brunâtre qui relie deux prolongements à la base des cuillères. Juste au-dessus de ce lien ténu, se dégage un passage rond, comme un trou, ménagé entre les deux cuillères : retenez le bien car c’est un passage secret déterminant dans le fonctionnement.
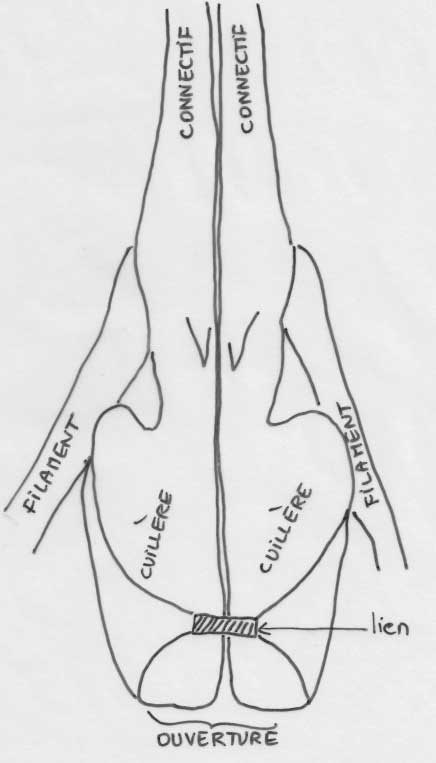
Schéma des deux étamines vues de face 
Les deux cuillères qui correspondent à la seconde loge stérile qui a migré vers la base 
Ouverture entre les deux cuillères réunies
L’étude du développement dans le bouton floral permet de comprendre comment se met en place cette structure complexe. Tôt dans le développement, deux étamines sur les quatre originelles avortent : elles sont réduites à l’état de minuscules staminodes quasi invisibles en haut du tube. Pour les deux autres, un développement asymétrique se met en place : le connectif qui relie les deux loges croît « anormalement » et les sépare ; l’une d’elles devient rapidement stérile et elle donnera le point d’attachement avec l’autre étamine. Côte à côte les deux longs connectifs parallèles et serrés ferment l’entrée de la corolle déjà bien rétrécie par les plis de la lèvre inférieure.

Fleur de sauge sclarée de face : entrée très rétrécie 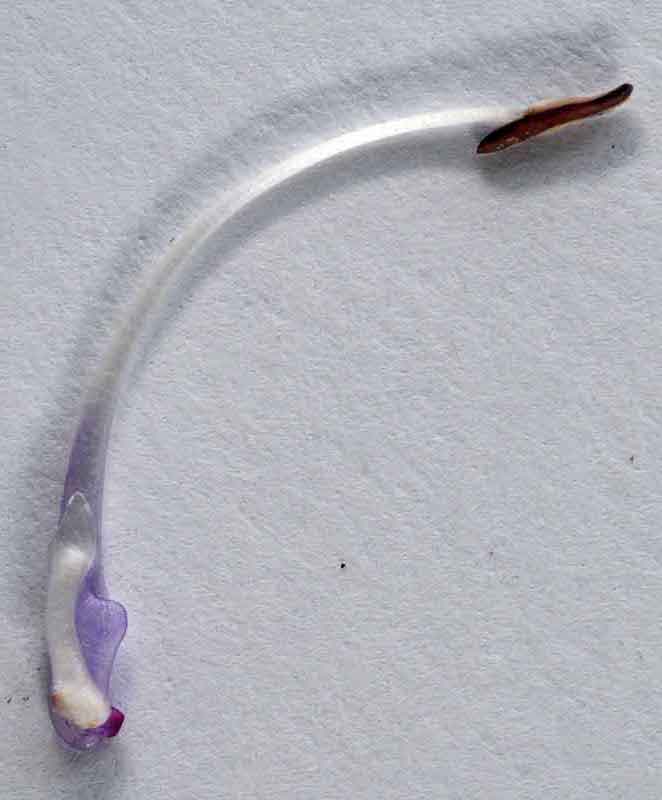
Etamine séparée avec sa cuillère basale moins développée que chez la sauge des prés 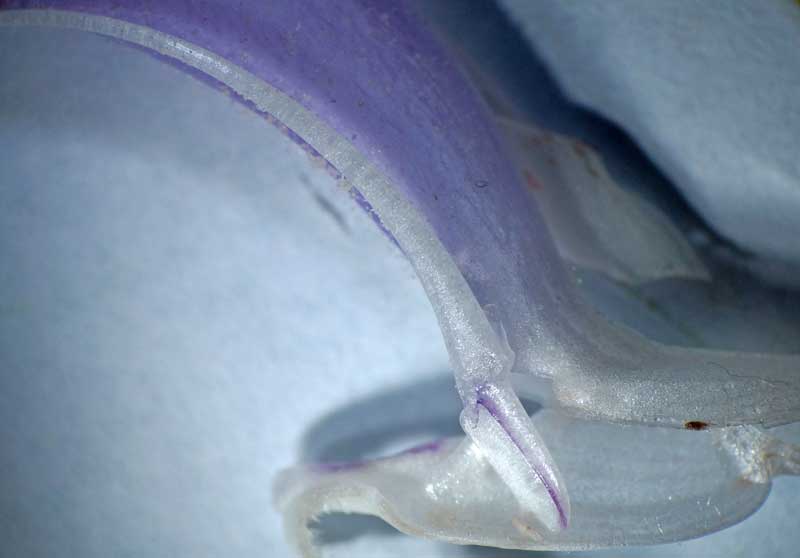
Articulation du filament sur le connectif
Mode d’emploi

Alors, comment çà marche ? Depuis la fin du 19ème siècle, on a propagé et médiatisé cet exemple de la sauge des prés d’une part comme archétype de toutes les sauges ce qui est faux (voir le dernier paragraphe) et d’autre part en le simplifiant à outrance. Ainsi dans l’ouvrage de référence de Proctor et Yeo paru en 1973 « The pollination of flowers », on peut lire : « Quand une abeille pousse sa tête dans la fleur, la plaque du connectif (la cuillère) est poussée vers le haut et en arrière et les lobes de l’anthère fertile basculent vers le bas apportant du pollen au contact de l’abdomen de l’abeille. » Cette vision dynamique du processus a été largement reprise dans les livres scolaires et les ouvrages de référence en biologie. Et pourtant, cette version s’avère tronquée et partiellement fausse au vu d’une étude récente. Effectivement, les deux cuillères et les filaments associés servent bien de bras de leviers permettant la bascule ; ils ferment l’entrée de la corolle tout comme les plis de la lèvre inférieure forçant l’abeille (le pollinisateur le plus fréquent chez la sauge des prés) et bloquent la tête de l’abeille : même les yeux restent visibles sur les cotés ; au plus les mandibules réussissent à passer dans goulot d’étranglement. Notons d’ailleurs que chez la sauge sclarée, l’entrée est encore plus étroite, la fleur semblant nettement comprimée latéralement vue de face ; par contre, la base des étamines n’a pas cet élargissement en cuillère typique de la sauge des prés !

Fleur de sauge sclarée de profil 
Fleurs de face : noter les plis de la lèvre inférieure et la gorge très étroite
Lors de cette butée, les étamines vont basculer rapidement et toucher peut-être le dos de l’abeille libérant une première dose de pollen. Mais le processus ne s’arrête pas là : l’abeille rapidement bloquée introduit alors sa langue par le fameux trou à la base des cuillères (voir paragraphe ci-dessus), détail clé complètement omis dans la version classique ! Pour atteindre au mieux le nectar tout au fond du tube, elle entreprend alors une seconde phase (très brève !) où elle appuie très fort avec sa tête ce qui accentue la pression sur la corolle et la base des étamines ; les forces exercées sur les cuillères entourant le trou où passe la langue font alors basculer l’étamine comme un bras de levier et la font appuyer encore plus sur le dos. En fait, les filaments articulés avec le reste de l’étamine se font à la fois pousser par la tête et tirer par les déformations de la corolle et tout ceci a lieu pendant une phase statique et non pas seulement pendant la première phase comme le suggère l’ancienne version ! L’angle avec lequel se positionne l’abeille, imposé par l’orientation de la lèvre inférieure s’avère déterminant tout comme la forte courbure du connectif de l’anthère fertile : si cette tige porteuse était droite, l’anthère ne toucherait pas le dos de l’abeille ! C’est donc toute l’architecture globale de la fleur qui intervient dans ce processus et pas seulement les leviers des étamines.

Distributeur de pollen
Au delà de déposer le pollen sur le dos de l’abeille, ce dispositif de levier présente au autre avantage majeur : il fonctionne à plusieurs reprises et n’est pas mis hors service au premier passage comme la projection explosive propre aux fleurs des papilionacées (voir la chronique avec les xylocopes). Comme la position du visiteur est très fortement contrainte par le dispositif d’ensemble (voir ci-dessus), alors les forces de frottement décisives entre les loges de l’anthère et le corps de l’abeille dépendent directement de l’étamine modifiée. La rigidité du connectif qui tient l’anthère s’avère ainsi déterminante : s’il est trop souple, l’anthère ne frottera pas assez et peu de pollen sera déposé durant la seconde phase statique ! De même sa courbure sera capitale pour pouvoir appuyer au maximum. Ainsi, de subtiles modulations de ce dispositif permettent de distribuer du pollen à chaque passage de visiteur. On estime qu’une fleur donnée distribue tout son pollen en 12 à 17 visites ce qui augmente fortement les chances de réussite.

Sauge hybride cultivée (S. X jamensis) 
Les deux étamines ferment complètement l’entrée 
Filament court et trapu portant le reste de l’étamine
Parmi les visiteurs des fleurs des sauges, un certain nombre tente de récolter du nectar mais se trouvent eux aussi bloqués par l’entrée rétrécie et souvent ne peuvent y accéder au nectar sauf s’ils disposent d’une langue assez longue. Par contre, du fait de leur moindre taille et faute d’un comportement adapté, ils ne réussissent pas à activer la bascule des leviers : la fleur ne gaspille pas son pollen ! Ainsi, chez la sauge sclarée, si les xylocopes semblent être les pollinisateurs clés et pratiquement les seuls capables d’actionner les leviers, on observe des visites d’abeilles solitaires « illégitimes » comme des mégachiles, des andrènes ou des syrphes. Les abeilles domestiques qui pollinisent activement la sauge des prés ne réussissent pas à atteindre le nectar de la sauge sclarée trop profond pour leur langue assez courte : elles essaient au plus de collecter du pollen directement sur l’anthère sous la lèvre supérieure. On voit donc se dessiner des différenciations fortes dans le cortège des pollinisateurs efficaces selon les espèces de sauges et la structure interne de leur fleur. On constate effectivement une forte diversification des agents pollinisateurs parmi les groupes de sauges ; ainsi, en Amérique, nombre d’espèces sont pollinisées par les colibris comme la sauge ananas cultivée comme ornementale ; elles partagent souvent des fleurs rouge intense, à lèvre supérieure non arquée et se tenant à l’horizontale.

4 exemples de « sauges à colibris » : la sauge ananas 
Sauge éclatante 
Sauge de Buchanan 
Sauge à involucre
Innovation réinventée

Sauge verticillée : des fleurs petites avec une lèvre supérieure peu différenciée 
Les étamines sont très simplifiées et le levier est pratiquement non fonctionnel 
Un bourdon en visite : les étamines ne basculent pas ; 25% des visiteurs sont des mouches !
Nous avons souligné en introduction « l’unité apparente » des sauges unies par le partage de l’élongation du connectif entre les deux loges des anthères contrairement aux autres genres de la tribu des Menthae. Dans le détail, on observe de nombreuses variations dans la morphologie de ces étamines basculantes mais avec, a priori, toujours la même structure de base ; d’où l’idée qui a longtemps dominé que les sauges étaient toutes étroitement apparentées et qu’elle dérivaient toutes d’un même ancêtre commun chez qui était apparu ce dispositif. Pourtant plusieurs analyses génétiques récentes intégrant diverses espèces de sauges et d’autres genres de la tribu montrent de manière récurrente et désormais admise qu’il existe trois lignées bien différenciées au sein des Sauges du genre Salvia telles qu’on les définit jusqu’alors. Si la disparition de deux étamines n’a eu lieu qu’une fois, l’évolution des deux étamines restantes s’est ensuite faite de manière différente. Cela veut dire qu’en fait le fameux dispositif des étamines à levier est apparu trois fois indépendamment au sein de la tribu des Menthae aboutissant à chaque fois à partir de structures légèrement différentes à un dispositif en apparence identique : bel exemple de convergence évolutive remarquable quasi impossible à détecter directement tant le résultats final se ressemble ! Ces études révèlent un autre fait encore plus troublant : chacune de ces lignées indépendantes de sauges se trouve plus étroitement apparentée à chaque fois avec un groupe de genres aux étamines non transformées qu’avec les deux autres lignées de sauges ! Ainsi, prenons le groupe où se trouvent la sauge sclarée et la sauge des prés, tout comme les sauges d’Ethiopie ou argentées : les parents les plus proches de ce groupe sont, non pas les autres sauges mais … les romarins et les Perovskias, arbustes ornementaux bien connus. Chez ce dernier, d’ailleurs surnommé (de manière prémonitrice ? !) sauge de Russie, il y a un très léger allongement du connectif entre les deux loges des anthères des deux étamines fertiles. Les étamines du romarin possèdent effectivement un connectif allongé dans leurs deux étamines avec une disparition complète de la loge stérile, soit un stade d’évolution digne de celui des sauges !

Fleurs de romarin officinal 
De profil, la ressemblance avec une fleur de sauge devient évidente ! 
Perovskia ou sauge de Russie 
Fleurs réduites et peu différenciées
Pour prendre en compte ces découvertes, on se dirige vers la conservation du genre Salvia tel que défini auparavant en distinguant à l’intérieur trois tribus dans lesquelles on intégrerait les genres apparentés comme les romarins qui deviendraient donc … des sauges !
Développement d’une étamine de sauge 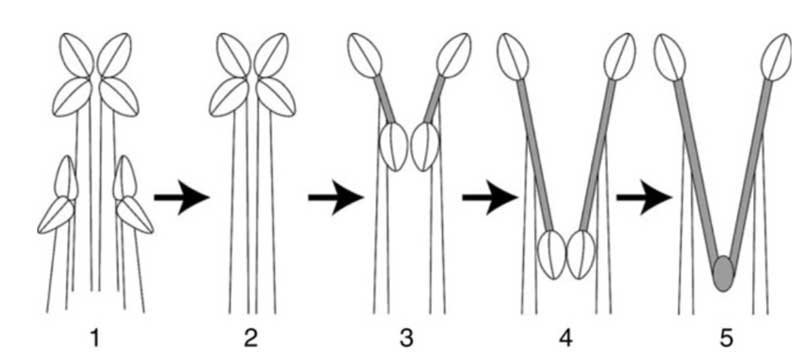
Scénario évolutif possible (en gris le connectif)
Enfin, on remarque à chaque fois que la lignée des sauges a développé de nombreuses espèces tandis que les genres apparentés ne contiennent que très peu d’espèces : l’acquisition d’étamines basculantes constitue donc une innovation évolutive clé, un « détonateur » qui booste les pressions de sélection et favorise la spéciation (voir l’exemple de l’éperon des ancolies).

Sauge fausse-germandrée 
Sauge farineuse 
Sauge bleu gentiane 
Sauge d’Ethiopie (espèce indigène malgré son nom !) 
Sauge officinale 
Sauge glutineuse (espèce montagnarde indigène)
BIBLIOGRAPHIE
The pollination and reproduction success of Salvia sclarea. Serdar Gökhan ŞENOL et al. Biological Diversity and Conservation 10/3 (2017) 130-135
New Insights into the Functional Morphology of the Lever Mechanism of Salvia pratensis (Lamiaceae).MARTIN REITH, GISELA BAUMANN, REGINE CLAßEN-BOCKHOFFand THOMAS SPECK. Annals of Botany 100: 393–400, 2007
The Staminal Lever Mechanism in Salvia L. ?Lamiaceae) a Review. R. Claûen-Bockhoff, P. Wester, and E. Tweraser Plant biol. 5 ; 2003) 33±41
Staminal Evolution in the Genus Salvia (Lamiaceae) : Molecular Phylogenetic Evidence for Multiple Origins of the Staminal Lever.JAY B. WALKER and KENNETH J. SYTSMA. Annals of Botany 100: 375–391, 2007
Salvia united: The greatest good for the greatest number Bryan T. Drew et al. TAXON 66 (1) • February 2017: 133–145
A retrouver dans nos ouvrages
Page(s) : 433-440

