Syrphes : des mouches expertes en pollinisation

Dans l’ordre des diptères qui réunit plus de 70 000 espèces de mouches, moustiques et moucherons, la famille des syrphes (syrphidés) se distingue à plus d’un titre : par son importance avec au moins 6000 espèces dans le monde dont plus de 500 en France ; par les colorations souvent vives et bigarrées de ces mouches et, pour nombre d’espèces, de fortes ressemblances mimétiques étonnantes avec des guêpes ou des abeilles ; par leur capacité à pratiquer le vol sur place (d’où leur nom populaire anglais de hoverflies) et pour certaines d’effectuer des migrations de masse sur de longues distances ; par la diversité des modes de vie de leurs larves (asticots en forme de limaces ou de vers) : saprophages (matériaux en décomposition), phytophages (herbivores), commensales (vivant dans des colonies d’insectes sociaux) ou prédatrices, mangeuses de pucerons très efficaces ; … Reste un dernier centre d’intérêt longtemps sous-estimé quant à son importance écologique et économique (par rapport à l’agriculture) : le rôle des syrphes adultes dans la pollinisation des plantes à fleurs ; les anglo-saxons ne les surnomment-ils pas flower flies, mouches des fleurs ? Tel sera donc le thème de cette chronique où nous allons découvrir que, dans ce domaine, les syrphes n’ont rien à envier aux abeilles et bourdons pourtant considérés comme les surdoués de la pollinisation par les insectes et, à ce titre, survalorisés par les médias.

Gros syrphe à allure de guêpe 
Les volucelles, très imposantes, peuvent rappeler des frelons !
Etre syrphe

Nous n’allons donc ici ne parler que des adultes seuls concernés par la pollinisation et leur portrait robot général. Au sein des diptères, les syrphes se situent dans l’ensemble informel des « mouches » ; d’ailleurs ce nom (masculin) de syrphe, apparu au tout début du 19ème siècle dérive du grec surphos qui signifie « mouche ». En tant que diptères, elles possèdent une seule paire d’ailes transparentes ce qui permet facilement de les distinguer, même pour les espèces très mimétiques, des guêpes et abeilles qui ont, elles, deux paires d’ailes transparentes (hyménoptères). Par rapport aux nombreuses autres « mouches », leurs ailes présentent deux traits caractéristiques (mais demandant un examen rapproché !) : une « fausse nervure » au milieu de l’aile (vena spuria) et un « faux bord » parallèlement à la bordure postérieure de l’aile !
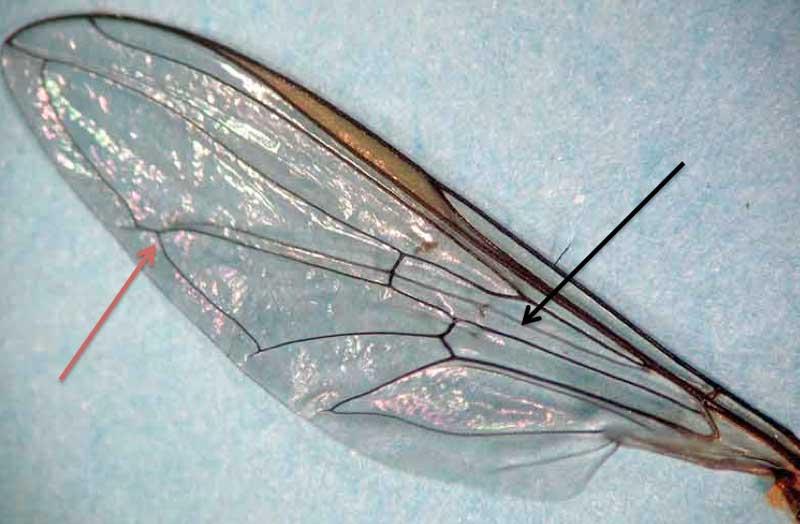
Aile de syrphe (flèche noire = fausse-nervure ; F. rouge = faux-bord) 
Aile de mouche bleue (non-syrphe)
La majorité des espèces ont des couleurs vives à base de jaune et de noir (souvent comme des guêpes) ou de teintes métalliques. La pilosité varie allant de limitée au seul thorax à recouvrant tout le corps comme chez les éristales mais on n’observe jamais de grandes soies dressées. On remarque vite la tête assez grosse, aussi large que le thorax, avec des yeux bien développés : le plus souvent, ces derniers sont très rapprochés chez les mâles et écartés chez les femelles. Les deux antennes courtes dirigées vers le bas (peu visibles !) surplombent souvent une fossette suivie d’un rebord, comme un « nez » au-dessus de la bouche avec les pièces buccales (voir ci-dessous). Taille et forme du corps varient énormément au sein de cette famille très diversifiée : de nombreuses espèces sont petites à moyennes (moins de 1cm) mais quelques genres atteignent les 2cm de long comme les grosses volucelles aux allures de bourdons. L’abdomen est souvent plus ou moins aplati et plutôt allongé. Terminons avec le trait comportemental le plus typique cité en introduction : le vol sur place, le plus souvent en plein soleil, avec de brusques décrochements pour aller se positionner de nouveau quelques mètres plus loin !

Trompe
Toutes les espèces de syrphes floricoles récoltent du pollen comme source principale de protéines ; ceci concerne tout particulièrement les femelles qui doivent en plus produire les œufs avec leurs réserves protéiques. Par contre, toutes ne prélèvent pas le nectar si la fleur en offre par ailleurs ; c’est le cas des genres de taille moyenne à petite avec des langue-trompes courtes même si d’autres genres de taille équivalente prélèvent à la fois nectar et pollen.

Syrphe en action sur un ibéris) 
Sur un fusain du Japon
Pour se nourrir, les syrphes, comme les autres mouches, disposent de pièces buccales transformées en appareil de type « suceur-lécheur » formé d’un rostre plus ou moins allongé d’où émerge une trompe. Cet ensemble varie considérablement en longueur selon les genres et espèces : généralement, il reste court autour de 2 à 4mm de longueur mais, chez les éristales et les volucelles plus grandes, il mesure de 5 à 8mm. Les rhingies, des syrphes assez communs, se distinguent par leur trompe très longue qui atteint 12mm, soit presque la longueur du corps quand elle est étendue : le rostre basal ainsi qu’une partie de la trompe (le labre) subissent un allongement important qui les font ressembler à des abeilles à langue développée !

Rhingie avec son rostre saillant 
Trompe déployée très longue mais terminée aussi par deux pièces (labelles)
La pointe de la trompe s’élargit en deux appendices ou labelles à la surface hérissée dont les syrphes se servent pour tapoter la fleur et recueillir le pollen superficiel. Ils peuvent aussi récolter le pollen en frottant les anthères des petites étamines entre les deux labelles ou, dans le cas d’anthères plus grandes en insérant un labelle à l’intérieur qui agit alors comme un racloir à pollen. Régulièrement, le syrphe nettoie sa face (souvent velue) et sa trompe avec ses pattes transférant ainsi les grains de pollen collectés vers la bouche pour le consommer. Les deux labelles peuvent aussi se rapprocher au point de former pour former comme un tube permettant d’aspirer le nectar des fleurs en tube ou au contraire s’étalent pour éponger le nectar étalé au fond des fleurs ouvertes. Cette diversité de modes d’emploi permet aux syrphes d’exploiter une large gamme de types de fleurs différents. Ainsi en Allemagne, on a recensé plus de 600 espèces de fleurs visitées par des syrphes et diverses études européennes montrent que jusqu’à 80% de la flore d’une région donnée peut être visitée par des syrphes !

La galerie photo ci-dessous montre quatre exemples différents de trompe repliée : la structure de base assez uniforme laisse néanmoins place à de subtiles variations dans la forme et l’ampleur des deux labelles terminaux.




Fleurs à syrphes

Syrphe aux côtés d’autres pollinisateurs « généralistes », des méligèthes (Coléoptères) 
Fleurs très ouvertes et simples de fusain du Japon 
Pollen très accessible sur ce Delosperma cultivé
En dépit de la diversité des espèces florales visitées, on peut néanmoins dégager un certain nombre de traits particuliers communs à celles-ci et les relier à l’anatomie et aux comportements des syrphes. Globalement, on constate logiquement qu’il existe une corrélation entre la longueur de la trompe, variable selon les espèces (voir ci-dessus), et la profondeur des corolles des fleurs visitées ; cependant, les espèces à longue trompe ne se privent pas pour autant d’exploiter aussi des fleurs très ouvertes et peu profondes en plus des autres ! Six grandes familles de plantes à fleurs dominent dans le paysage floral des syrphes : les ombellifères (ou apiacées), les composées (ou astéracées), les renonculacées, les crucifères (ou brassicacées), les caryophyllacées et les rosacées. Or, ces familles correspondent justement dans leur majorité à des espèces de fleurs dites généralistes, i.e. visitées par un grand nombre d’espèces d’insectes appartenant à des groupes très différents et pour la plupart peu spécialisés. De plus, on note une forte propension de ces mouches à visiter des fleurs dites anémophiles (i.e. pollinisées par le vent), réputées a priori comme n’intéressant pas les pollinisateurs : des graminées ou des plantains par exemple ; elles y trouvent du pollen en grande quantité et ne sont pas perturbées par l’absence de nectar.

Fleur de compagnon blanc (Caryophyllacée) 
Coquelicot (Papaveracée) 
Peucédan officinal (Apiacée) 
Grande berce (Apiacée) 
Aubépine (Rosacée) 
Rose cultivée (Rosacée)
Les espèces dotées d’une trompe plus longue, peuvent accéder à une gamme plus large de fleurs dont des familles florales plus spécialisées. Ainsi, ces syrphes butinent les petites fleurs irrégulières avec un tube et des lèvres des labiées (ou lamiacées) ou des scrophulariacées : épiaires, bugles, lierre terrestre ; elles savent s’en approcher de manière appropriée pour atterrir et gèrent très facilement l’accès au pollen et au nectar cachés au sein de la corolle et moins accessibles. Des mouches classiques comme les mouches à viande vertes ou les mouches domestiques, elles aussi floricoles, ne visitent de telles fleurs que par hasard et sans réussir à atteindre leurs ressources. De même, ces syrphes à trompe moyenne recherchent les petites fleurs des papilionacées (ou fabacées) comme les mélilots, les luzernes ou les trèfles, elles aussi plus difficiles à gérer du fait de la corolle irrégulière. Citons aussi une partie de la famille des caprifoliacées avec les scabieuses, les knauties et la succise (voir la chronique sur cette dernière espèce), aux fleurs un peu irrégulières.

Knautie 
Succise
Syndrome syrphal ?

Il ressort jusqu’ici une forte impression de généralisme de la part des syrphes. Pourtant, il semblerait que quelques espèces de notre flore aient un rapport étroit et plus spécialisé avec les syrphes : ce sont quelques espèces de véroniques forestières (comme la véronique officinale ; voir aussi la chronique sur la véronique de Perse) et les circées (voir la chronique sur ces plantes). Leurs fleurs petites, peu différenciées et réduites partagent une disposition très particulière de leur unique paire d’étamines (un caractère déjà peu courant) : elles sont étalées sur les côtés avec leurs anthères chargées de pollen dirigées vers le bas et disposées à un niveau légèrement plus bas que le stigmate central dressé (l’organe du pistil récepteur des grains de pollen). Les syrphes de petite taille qui visitent ces fleurs agrippent les anthères pour prélever le pollen ce qui provoque un affaissement des étamines sous leur poids ; ainsi, le dessous de leur corps un peu velu et éventuellement porteur de grains de pollen accrochés au passage entre en contact avec le stigmate et assure ainsi la pollinisation croisée. Par ailleurs, les plantes concernées habitent des milieux boisés humides et ombragés propices comme habitat pour le cycle de vie des petites espèces de syrphes (Melanostoma, Baccha, Syritta) alors que les abeilles solitaires de taille équivalente susceptibles de les butiner y sont plutôt rares et peu actives.

Fleurs de véronique officinale 
Fleurs de circée de Paris
A l’autre extrême, les syrphes à très longues trompes comme les rhingies tendent à se spécialiser sur des fleurs plus complexes en tube à la manière des abeilles et bourdons à longue langue : les compagnons rouges, les jacinthes des bois, les pulmonaires, les myosotis, les balsamines à petites fleurs et les liserons. Ainsi, les rhingies recherchent activement les liserons des haies aux grosses fleurs blanches profondes et viennent souvent jusque dans les jardins exploiter des espèces ornementales exotiques comme les ipomées.
Mais la spécialisation la plus extrême se trouve atteinte avec certaines espèces d’orchidées exotiques. Ainsi, au brésil, l’orchidée Govenia utriculata est exclusivement visitée par deux espèces de syrphes locales : la fleur présente des petits points jaune à orange et brunâtres qui imitent des paquets de pollen tombé et attirent les syrphes ; quand elles cherchent à récolter ce « faux-pollen » leur tête heurte la base des organes reproducteurs et le vrai pollen se colle sur leur tête, hors d’atteinte pour elles. Encore plus surprenant le cas d’une orchidée du Moyen-Orient (jusqu’à Chypre) : l’épipactis à feuilles de vératre. Elle est exclusivement pollinisée par cinq espèces de syrphes dont au moins trois très communes comme le syrphe ceinturé ou le syrphe porte-plume. Les fleurs produisent des substances volatiles spécifiques qui induisent le comportement de ponte des syrphes : ce faisant, elles prennent en charge le pollen qui se colle sur elles. L’analyse de ces composés chimiques émis par l’orchidée montre qu’ils sont très proches des phéromones d’alarme émises par certaines espèces de pucerons quand ils se sentent menacés ; comme ces syrphes ont des larves prédatrices des pucerons, elles pondent sur des plantes en les croyant porteuses de pucerons !

Irrésistible jaune
On constate que parmi les nombreuses fleurs visitées, la couleur jaune domine largement : chez les brassicacées (dont le colza pour lequel les syrphes sont des pollinisateurs majeurs), chez les renonculacées avec les boutons d’or notamment et surtout chez les astéracées avec les épervières, les liondents, picrides, pissenlits, laiterons, … aux capitules jaunes très visités.

Onagre 
Doronic 
Molène 
Pavot de Californie
Une étude très fine a exploré cette préférence apparente chez l’éristale gluant, un syrphe très velu, très commun et ressemblant à une abeille domestique. Des expériences ont cherché à déterminer ce qui déterminait le comportement d’atterrissage sur une fleur donnée et, ensuite, l’extension de la trompe pour prélever le pollen et le nectar. Les résultats démontrent clairement une préférence innée pour les couleurs jaunes ; or, cette couleur se trouve souvent associée à des phénomènes de réflexion des ultra-violets (UV). Par des expériences de choix multiples avec des fleurs artificielles, on a testé des éristales inexpérimentés et naïfs (juste éclos). Pour atterrir, le fait que le jaune absorbe ou reflète les UV ne semble pas déterminant ; elles se posent même sur des fleurs artificielles non jaunes mais brillantes et absorbant fortement les UV. Une fois posées, ils étendent leur trompe de préférence vers des taches jaune sombre ou absorbant les UV. La luminosité a donc une importance dans le choix des syrphes et, probablement, que les préférences dépendent du type de réflexion des UV induit par la couleur jaune.
Cet attrait pour le jaune a aussi été démontré avec un autre exemple de relation spécifique envers une orchidée exotique, un sabot de Vénus du sud-est asiatique (Paphiopedilum barbigerum) aux fleurs visitées par deux espèces de syrphes (dont l’universel syrphe ceinturé !). Ils atterrissent sur un appendice charnu jaune et glissant (staminode) qui les fait tomber dans la poche en sabot où ils restent prisonniers un instant, assurant au passage le transfert du pollen collé sur leur corps. Or, cet organe reflète fortement la couleur jaune appréciée des syrphes et il n’y a pas de substances volatiles émises.
Grande constance
Les syrphes s’avèrent être de bons pollinisateurs à cause d’un trait comportemental récurrent : la fidélité florale. Ainsi, des analyses de pollen présent dans le tube digestif de syrphes du genre Melanostoma, espèces généralistes, ne révèlent que moins de 3 sortes de pollens différents alors que l’environnement immédiat offre des dizaines ou des centaines d’espèces florales potentiellement visitables. Cette fidélité individuelle à quelques espèces était bien connue chez bourdons, abeilles ou papillons mais n’a été démontrée que récemment chez les syrphes, longtemps sous-estimées du fait de leur statut de « mouches ».
Deux espèces communes, le syrphe ceinturé et le syrphe des groseilliers, ont été testées expérimentalement en leur présentant deux variantes colorées d’une même espèce très appréciée, la lobulaire maritime (voir la chronique sur les vergers bio) sur laquelle on les laisse butiner ; ensuite, on place cette plante au milieu de parterres variés. Les deux syrphes montrent alors une forte constance envers cette espèce mais sans faire de distinction envers le morphe coloré sur lequel elles ont été entraînées au départ. Les deux variantes offrent autant de pollen et aussi accessible dans les deux cas.
Pour expliquer cette fidélité florale, on a souvent invoqué une meilleure efficacité dans le traitement des fleurs et de leurs ressources mais ici il s’agit d’une fleur simple où ils ne prennent que le pollen très facile d’accès. On a aussi avancé la formation d’une « image de recherche » par mémorisation mais ici les syrphes ne semblent pas mémoriser la forme colorée. Peut-être que leurs récepteurs visuels ne leur permettent pas de discerner les fleurs blanches des fleurs violettes, ces derniers étant avant tout réglés sur le jaune (voir ci-dessus). En tout cas, ils se montrent bel et bien capables « d’apprendre » très vite à la manière des abeilles et bourdons.
Méthodique
Le grand spécialiste des syrphes, F.S. Gilbert, dans les années 1980, a mené d’innombrables études expérimentales sur les syrphes avec une précision des détails qui laisse pantois (typiquement « british » !). Il a ainsi cherché à comprendre la méthode des syrphes pour traiter les capitules de composées formés de nombreuses fleurs élémentaires ou fleurons et constituant chacun une « tache » de ressource très dense : il a observé minutieusement des éristales gluants en train de butiner des capitules d’aster américain en notant le temps passé, le nombre de fleurons sondés, l’angle et la distance de départ jusqu’au capitule suivant, les quantités de nectar secrétées par les fleurons, …
Pour comprendre ce qui se passe, il faut d’abord bien appréhender la complexité de la structure des capitules formés de centaines de fleurons élémentaires disposés selon des spirales entrelacées (voir la chronique La reine des anneaux fleuris au paragraphe sur les spirales) ; mais, du point de vue des visiteurs, l’important concerne le processus de maturation et de floraison des fleurons. Ceux-ci mûrissent selon des cercles et la floraison progresse de l’extérieur vers le centre ; ainsi, à tout moment sur un capitule, on a un centre formé de fleurons immatures encore fermés, puis un cercle unique de fleurons dont les anthères mûres libèrent du pollen, puis quelques cercles au stade femelle avec les stigmates sorties et enfin les cercles extérieurs déjà passés et fanés brunis. Par ailleurs, le pollen se trouve poussé par un système de piston exercé par le stigmate ascendant (comme chez les proches campanulacées : voir la chronique) et présenté en amas au sommet du fleuron.

Quand un éristale arrive sur un capitule d’aster, il exploite le cercle de fleurons porteurs de pollen et l’ingère en se nettoyant les pattes et la face ; méthodiquement, fleuron après fleuron, il fait le tour du cercle (en en sautant quand même quelques uns) pour sonder la présence de pollen et nectar puis dans 95% des cas, s’envole vers un autre capitule. La visite d’un cercle qui compte en moyenne 19 fleurons lui prend environ 20 secondes. Mais comment fait-il pour savoir de manière aussi précise qu’il a fait le tour à 360° ? Il ne peut pas évaluer la quantité de nectar prélevé car seulement 10% des fleurons en contiennent et en très petites quantités ; le chercheur s’est « amusé » à tourner délicatement un capitule à contre courant du mode opératoire de l’éristale qui, dans ce cas, peut sonder quatre fois de suite les mêmes fleurons, preuve que la quantité prélevée n’intervient pas. Il doit se positionner par rapport à la lumière et plus particulièrement au plan de polarisation de celle-ci, un facteur auxquels ils sont sensibles comme les abeilles. Il se peut aussi qu’ils aient une mémoire spatiale très fine de repères visuels comme cela a été démontré par ailleurs. On voit donc qu’en termes de capacités les syrphes n’ont rien à envier aux abeilles !
Mouches-abeilles

Il se dégage donc que les syrphes, fonctionnellement, se montrent souvent égaux aux hyménoptères classiquement considérés comme les pollinisateurs incontournables et inégalables en matière d’efficacité. Ils possèdent des attributs sensoriels et comportementaux équivalents dont la capacité de mémorisation et de fidélisation. De fait, certains syrphes se montrent capables de manipuler des fleurs à l’architecture complexe (corolle irrégulière) avec une grande aisance ; si on ajoute leur capacité de fidélisation à l’échelle individuelle, tout concourt pour en faire de très bons pollinisateurs. Effectivement, on découvre que souvent des fleurs dites « à abeilles » se trouvent en fait aussi pollinisées par des syrphes dans des régions ou à des périodes où les abeilles et bourdons sont rares ou absents. Nous avons vu le cas des espèces de véroniques et circées des bois humides ombragés et cela pourrait s’étendre à d’autres environnements.

Avec le fort déclin des abeilles et bourdons lié au changement global, on commence à observer une augmentation du nombre de syrphes, au moins les espèces ubiquistes et capables de vivre dans des milieux ouverts et transformés comme le syrphe ceinturé. Ceci signifie qu’il va falloir vraiment prendre en compte les syrphes comme agents de pollinisation et notamment repenser les aménagements en zones agricoles (comme les bandes fleuries ou jachères), conçus avant tout par rapport aux abeilles et bourdons… Sans oublier que nombre de ses espèces constituent de plus de précieux axillaires pour le contrôle biologique des pucerons via leurs larves prédatrices : avec les syrphes, c’est le deux en un !
Bibliographie
Pollination and floral ecology. P. Willmer. Princeton university Press. 2011
The Yellow Specialist: Colour Preferences and Colour Learning of the Hoverfly Eristalis tenax (Diptera: Syrphidae) ALEXANDER NEIMANN, LINA AN & KLAUS LUNAU. Entomologie heute 30 (2018): 27-44
Pollination by deceit in Paphiopedilum barbigerum (Orchidaceae): a staminode exploits the innate colour preferences of hoverflies (Syrphidae). J. Shi et al. Plant Biology a 2008
Smells like aphids: orchid flowers mimic aphid alarm pheromones to attract hoverflies for pollination. Johannes Stökl et al. Proc. R. Soc. B (2011) 278, 1216–1222
Reproductive biology and pollination of Govenia utriculata: A syrphid fly orchid pollinated through a pollen-deceptive mechanism. EMERSON R. PANSARIN. Plant Species Biology (2008) 23, 90–96
The foraging ecology of hoverflies : circular movements on composite flowers. F.S. Gilbert. Behav. Ecol. Sociobiol. (1983) 13 : 253-257
Flower constancy in the hoverflies Episyrphus balteatus (Degeer) and Syrphus ribesii (L.) (Syrphidae). Dave Goulson and Nick P. Wright Behavioral Ecology 1997 Vol. 9 No. 3: 215-219
