La colonisation du sous-bois des jeunes forêts
A propos des forêts françaises, deux évènements ont récemment été médiatisés : la plantation d’une forêt « primitive » dans le pelouse de Reuilly du Bois de Vincennes (2000 arbres plantés de 23 espèces différentes) et, plus étonnant, le début de la plantation d’une forêt de … 1350 hectares dans la plaine de Pierrelaye-Bessancourt (val d’Oise) sur des terres agricoles polluées par l’épandage d’eaux usées. Désormais, on plante surtout des feuillus, en variant les essences et en privilégiant des essences indigènes, deux conditions connues pour favoriser la restauration de la biodiversité associée à ces forêts nouvelles, notamment la flore herbacée des sous-bois. Mais celle-ci se réinstalle-t-elle vraiment et à quelle vitesse ? Une étude suédoise portant sur des terres agricoles abandonnées récemment, jouxtant des massifs forestiers, plantées en feuillus, apporte des éléments très intéressants sur cette colonisation par les plantes herbacées du sous-bois et casse quelques idées reçues sur la vitesse du processus.

Référence biaisée
La vitesse de colonisation du sous-bois des forêts nouvelles par les espèces herbacées depuis la forêt adjacente dépend de divers facteurs écologiques : l’éloignement par apport à la forêt ancienne source de graines/fruits ou spores pour la dispersion ce qui prend en compte la taille de la parcelle reboisée (périphérie versus cœur) ; leur tolérance à l’ombre dispensée par la canopée des arbres installés (même dans une jeune forêt). Les traits d’histoire de vie des espèces concernées jouent un rôle décisif dans ce processus de colonisation : la masse de leurs fruits/graines, la forme de celles-ci et leurs adaptations à tel ou tel mode de dispersion par exemple par le vent ou anémochorie (voir les chroniques sur ce thème), par accrochage au pelage des animaux (voir les chroniques sur ce thème) ou par les fourmis (myrmécochorie : voir l’exemple de l’hellébore fétide) ; leur capacité à s’étaler sur place via la multiplication végétative par rhizomes (voir l’exemple de l’oyat) ou par stolons (voir l’exemple du fraisier) ; ou encore leur capacité à résister à la compétition au sein de la strate herbacée via leur potentiel de croissance.
Les études précédentes sur cette colonisation dans d’anciennes parcelles agricoles reboisées en bordure de forêts établies convergeaient vers un constat admis comme vérité générale : la majorité des espèces forestières herbacées du sous-bois avaient une vitesse de colonisation très basse, rarement au-dessus de 2,5m par an ! Cependant, ces études portaient sur des parcelles de petite taille (moins de deux hectares) et déjà relativement « anciennes » avec un âge entre 30 et 100 ans : deux conditions réductrices qui biaisaient peut-être les résultats obtenus ? C’est pourquoi cette nouvelle étude a été entreprise en Suède sur d’anciennes parcelles agricoles reboisées en feuillus de taille variée (des grandes et des petites) et très jeunes (entre 14 et 18 ans) : ainsi, on se trouve dans des situations plus proches d’une colonisation réelle en cours. Chaque parcelle reboisée est comparée avec la forêt adjacente.
Double erreur

Jusqu’ici, on tendait à considérer que toutes les espèces herbacées des sous-bois tolérantes à l’ombre de la canopée formaient un groupe homogène avec une faible capacité de colonisation. Or, les valeurs de vitesse maximale de colonisation obtenues ici s’avèrent être bien plus conséquentes que celles admises jusqu’ici comme le montre le tableau d’exemples ci-dessous.
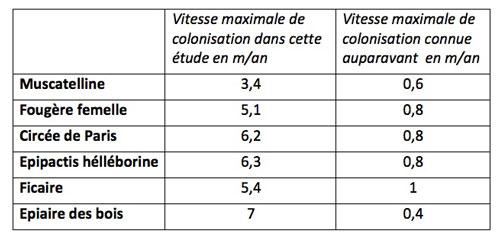
On trouve ici des valeurs 8 à 20 fois plus élevées que celles admises jusqu’ici. Même si ces valeurs maximales peuvent sembler modestes, à l’échelle d’une vingtaine d’années cela change beaucoup de choses dans la colonisation d’un sous-bois : le front de progression va avancer bien plus vite. Il augmente de plus la probabilité qu’il y ait de rares évènements de dispersion à longue distance qui vont faire bondir le front loin en avant pour les espèces concernées. Au cours de leur étude qui portait donc sur des parcelles très jeunes (moins de 20 ans), les chercheurs ont pu constater qu’un certain nombre d’espèces avaient effectivement déjà établi des belles populations montrant qu’elles n’étaient pas limitées par leur capacité de dispersion.

Le second enseignement de cette étude est que le groupe des colonisatrices n’est pas du tout homogène quant à cette capacité de colonisation : au contraire, on peut distinguer deux grands groupes bien distincts : les colonisateurs rapides et les colonisateurs lents. Ils diffèrent par leurs traits d’histoire de vie : chacun de ces deux sous-groupes possède une combinaison propre de ces traits comme nous allons le voir.
Les lièvres
La vitesse de colonisation augmente fortement avec la hauteur des plantes et leurs distances naturelles de dispersion ; par contre, le poids des graines/fruits et la capacité à s’étaler sur place par multiplication végétative sont peu corrélés avec cette vitesse de colonisation, voire avec un effet négatif. Le groupe des « lièvres » se compose donc globalement de plantes relativement grandes, ne recourant pas à la multiplication végétative et avec des fruits/graines qui s’accrochent aux animaux ou transportés par le vent. Le tableau ci-dessous en donne une quinzaine d’exemples.

Pâturin des bois 
Chiendent des bois 
Fétuque géante 
Brachypode des bois 
Laîche des bois 
Ail des ours 
Epiaire des bois 
Scrofulaire noueuse
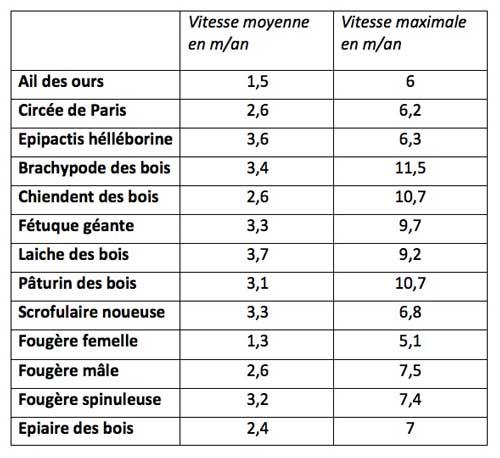
Des plantes plus hautes ont effectivement plus de chances de voir leurs fruits/graines s’accrocher à des animaux de passage qui sont ici essentiellement des mammifères de taille moyenne : lièvres, renards, sangliers et chevreuils ; les espèces les plus grandes ont un rayon d’action encore plus important et peuvent donc emporter les fruits/graines accrochés à grande distance (voir la chronique sur l’exemple des sangliers). Donc plus la plante est grande, plus elle a de chances de « croiser » un grand animal. Ceci explique que les plantes avec des fruits/graines assez gros ne semblent pas pour autant handicapées dès lors qu’elles ont un dispositif d’accrochage efficace. D’autre part, des fruits/graines plus en hauteur ont aussi plus de chances d’être emportés par les rafales de vent (atténuées en sous-bois) pour les espèces anémochores (à graines/fruits transportées par le vent).

Circée de Paris 
Fruits accrocheurs de la circée
Parmi les lièvres figurent aussi des fougères et une orchidée (épipactis héléborine) : elles partagent le fait de se propager par des spores ou des graines minuscules, très légères et donc transportées à grande distance. Une étude précédente avait d’ailleurs démontré que dans le cas de la fougère spinuleuse, l’installation n’était pas limitée par la dispersion mais par la disponibilité de microsites favorables comme des vieilles souches pourries, de suintements humides, …

Fougère femelle 
Fougère mâle 
Fougère spinuleuse
Les tortues

Sabline à trois nervures 
Luzule poilue 
Sceua de Salomon multiflore
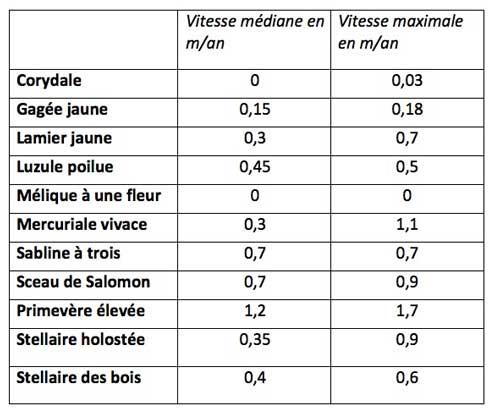
Le groupe des colonisateurs lents renferme de son côté des espèces plutôt basses, pratiquant souvent la multiplication végétative (rhizomes, stolons, bulbilles, …) et se dispersant par des fruits/graines assez gros soit via les fourmis avec la présence d’appendices huileux (élaïsomes), soit sans véritable dispositif particulier de dispersion. La comparaison des espèces dispersées par les fourmis avec celles dispersées par le vent confirme la faible capacité de dispersion des premières, les fourmis ne déplaçant souvent les graines que de un mètre au plus.

Le tableau ci-dessus en donne divers exemples.

Parmi ces tortues, certaines peuvent montrer une vitesse très lente au début de la colonisation dans les très jeunes forêts puis voir ensuite cette vitesse augmenter sensiblement avec le vieillissement du peuplement : c’est le cas d’espèces à floraison printanière (vernales) telles que les corydales ou la gagée jaune.

Corydales au printemps 
Gagées jaunes en mai
Parmi le cortège des espèces vernales, l’anémone sylvie (voir la chronique sur cette espèce) domine et avance régulièrement selon un front de colonisation ; l’absence de fourmis comme agents disperseurs dans nombre de très jeunes forêts peut expliquer son installation plus ou moins tardive.

Stellaire holostée 
Stellaire des bois
Par contre, dans ce groupe, les espèces dites clonales (voir l’exemple du muguet), i.e. capables de se multiplier végétativement, réussissent quand même à progresser d’environ 1m/an par expansion sur place, de proche en proche ; cette expansion ne dépend aps de la présence d’agents de dispersion puisque c’est la plante qui fabrique ses stolons ou rhizomes. Le lamier jaune, la stellaire holostée et la stellaire des bois en sont de bons exemples.

Rémanences
Le sol joue un rôle tout aussi déterminant dans ce processus de colonisation. Or, les très jeunes forêts étudiées ont été plantées sur d’anciennes terres agricoles qui ont reçu sur de longues périodes des engrais avec notamment du phosphore qui persiste ensuite très longtemps (peu lessivé). Les sols de ces jeunes forêts se trouvent donc plus riches en nutriments essentiels dont le phosphore ; or, on constate que les petites espèces de sous-bois apprécient bien l’enrichissement en phosphore mais doivent faire face à la compétition inégale avec de grandes plantes généralistes telles que la grande ortie, très envahissante via ses rhizomes. Les espèces plus grandes et avec un taux de croissance élevé seront plus aptes à lutter contre l’ortie. Par contre, ces ex-terres agricoles possèdent souvent un contenu en matière organique réduit ce qui peut au moins dans les premières décennies limiter l’installation et l’expansion de petites espèces liées à la présence d’humus riche telles que l’oxalis petite-oseille, la mélique uniflore ou l’aspérule odorante.

Oxalis petite-oseille 
Mélique uniflore (Graminée)
Ces différences de composition de sol agissent aussi sur la croissance des jeunes arbres (dans cette étude, des chênes et des bouleaux) qui peuvent développer ainsi une canopée plus dense entravant l’installation des herbacées de sous-bois même si elles tolèrent un certain ombrage.

Espoir

Divers exemples montrent que les plantations de feuillus peuvent ainsi contribuer à l’installation d’une riche biodiversité forestière quand elles sont installées sur des sols dégradés, si on replante des espèces indigènes et si la plantation jouxte un massif forestier (ou bien si on créé des corridors forestiers assurant des connexions). Dans l’exemple suédois étudié, plusieurs espèces forestières rares classées sur la liste rouge des espèces menacées en Suède ont été trouvées dans ces jeunes boisements : la muscatelline, la primevère élevée ou la véronique des montagnes.

Véronique des montagnes 
Muscatelline
Une autre raison d’être optimiste concerne l’évolution des sols de ces nouvelles forêts. Dans les forêts anciennes, le sol tend à s’acidifier naturellement et on a surtout déforesté des forêts aux sols riches et basiques pour en faire des cultures : ceci limite le maintien ou la présence de nombreuses espèces recherchant des sols plutôt neutres ou basiques, lesquelles connaissent justement un net déclin historique. Or, ici, il a été démontré que dans des chênaies installées sur d’anciennes terres agricoles, le sol reste basique (du fait des amendements passés) bien plus longtemps (jusqu’à au moins 80 ans) ce qui favorise donc l’installation de ces espèces qui, en pus, font majoritairement partie des tortues !

Bibliographie
Life-history traits explain rapid colonization of young post-agricultural forests by understory herbs.Jörg Brunet, Pieter De Frenne, Emma Holmström, Marian Lajos Mayr. Forest Ecology and Management 278 (2012) 55–62
A retrouver dans nos ouvrages
Page(s) :

