La fragmentation forestière transforme les hêtres
Fagus sylvatica

Parmi les nombreux grands problèmes écologiques qui affectent la biodiversité, la fragmentation des habitats figure en « bonne » place : de nombreux habitats dont les forêts ont subi et subissent les coups de boutoir de la déforestation et des aménagements divers (urbanisation, routes et autoroutes, …) qui transforment des massifs forestiers continus en fragments ou ilots séparés, plus ou moins isolés les uns des autres. Cette fragmentation modifie évidemment les conditions écologiques qui régnaient dans le milieu continu en introduisant notamment des effets de lisière majeurs et en facilitant l’action d’autres processus comme la pénétration humaine, les contacts avec des pollutions environnantes, l’entrée des espèces envahissantes, la pression de prédation, … (voir les autres chroniques consacrées à la fragmentation). Mais, il existe un autre ensemble d’effets bien plus insidieux parce que très durs à détecter et à démontrer et qui agissent souvent à moyen voire à long terme : les effets sur la génétique des populations des espèces subsistant dans ces fragments avec entre autres les conséquences de la consanguinité et/ou de la dérive génétique. Or, la génétique reste déterminante pour le succès reproductif de celles-ci puisqu’elle se trouve au centre du processus de la sélection naturelle qui agit sur les mutations. Avec la crise climatique en cours et qui s’accentue, les populations locales des fragments isolés vont avoir plus que jamais besoin de disposer d’un fort potentiel de variations génétiques en leur sein pour réussir à surmonter le terrible challenge qu’elles affrontent et se maintenir en s’adaptant. On avait tendance à croire jusqu’ici que les arbres forestiers en général avaient vis-à-vis de ces problèmes génétiques une forte capacité de résistance : une étude sur les hêtres dans un contexte de fragmentation forestière ancienne démontre clairement le contraire !
Anémophile
Dès lors qu’on s’intéresse à la génétique des populations d’une espèce d’arbre, il faut aborder deux aspects de sa biologie : d’une part son mode de reproduction avec notamment la pollinisation et d’autre part son mode de dispersion de ses graines. Ces deux évènements déterminent la circulation des gènes, le flux de gènes comme disent les biologistes, entre individus qui oriente l’évolution génétique de la population sous la pression de la sélection naturelle.

Chatons de fleurs mâles 
Fleurs femelles fécondées et chatons mâles 
Futur faux-fruit issu de la fécondation des fleurs femelles enfermées dans la cupule « épineuse »
Le hêtre (Fagus sylvatica), comme les autres membres de la famille des Fagacées (voir la chronique sur les fagales) a un système reproducteur à sexes séparés mais réunis sur la même plante individuelle : on parle d’espèce monoïque. Les hêtres ne commencent à se reproduire qu’à partir de l’âge de 40 ans. Les fleurs mâles avec leurs étamines qui produisent le pollen sont réunies en petites boules globuleuses (des chatons) au bout de longs pédoncules à la base des jeunes rameaux de l’année ; les fleurs femelles, porteuses du pistil avec l’ovaire contenant les ovules (les futures graines), sont elles groupées par deux à quatre dans une petite cupule verte vaguement armée de pointes molles et pendent au bout des rameaux. Cette floraison, qui a lieu en avril mai alors que le feuillage est déjà bien déployé, passe très facilement inaperçue, surtout que les chatons se trouvent souvent en hauteur dans la canopée. Les chatons mâles libèrent le pollen entraîné par le vent : on qualifie le hêtre d’espèce anémophile ; les fleurs femelles captent le pollen à l’aide de leurs stigmates qui dépassent.
Pour que la fécondation ait lieu, il faut que le pollen intercepté par les fleurs femelles soit reconnu et accepté et puisse germer sur les stigmates. Or, chez le hêtre, cette reconnaissance écarte très fortement (mais pas entièrement) le pollen issu du même arbre qui porte les fleurs femelles, via un système de reconnaissance génétique à la manière du système immunitaire (autoincompatible). Ceci prévient donc largement l’autofécondation, source de problèmes génétiques à court et moyen terme.

Zoochore
Les fleurs femelles fécondées par des grains de pollen interceptés se transforment en « faux » fruit : la cupule externe grandit et se durcit donnant comme une « bogue » un peu épineuse et qui renferme de deux à quatre « vrais » fruits, les faînes dures et triangulaires. Chaque fleur femelle fécondée donne un fruit, donc une faîne. Chaque faîne renferme une seule graine qui occupe pratiquement tout le volume interne et dont la paroi est soudée à celle de la faîne. C’est pourquoi on a l’impression que la faîne est une graine : c’est une graine-fruit ou akène.

Faux-fruit en cours de maturation 
A maturité, les coques ligneuses s’ouvrent en quatre et laissent tomber les faînes 
Faux-fruit ouvert avec deux faînes (les vrais fruits) 
Chaque faîne renferme une seule graine
En général, il y a deux faînes par faux-fruit : la cupule à maturité, devenue dure et ligneuse, s’ouvre en quatre valves qui exposent les faînes d’une belle couleur noisette et comestibles. Si elles ne sont pas récoltées et mangées par divers prédateurs dont des oiseaux comme les mésanges charbonnières ou les sitelles, elles finissent par tomber au sol au pied de l’arbre qui les a produites. Si elles restent là, elles pourront germer ensuite mais les plantules qui émergeront n’auront pratiquement aucune chance de se développer sous l’ombre très forte et dense de leur géniteur et avec la concurrence du réseau de racines déployé juste en dessous du sol.

Faux-fruits et faînes tombent au sol sous l’arbre producteur 
Germinations de plantules : les deux cotylédons ne ressemblent pas du tout aux feuilles adultes ! 
Jeunes plantules avec leurs deux premières vraies feuilles au-dessus des cotylédons
Pour avoir des chances de « réussir » en donnant un nouvel arbre capable de se développer, les faînes doivent être transportées hors de la zone d’influence de la canopée et de l’appareil racinaire. Autrement dit, elles doivent bénéficier d’une seconde dispersion, après celle qui les a simplement jetée à terre, avec l’aide involontaire de petits animaux tels que les mulots ou les écureuils qui, certes en mangent une partie (donc elles sont perdues pour l’arbre) mais en cachent une autre partie au sol sous les feuilles mortes, dans des crevasses de souches, sous du bois mort, … plus ou moins loin de là où ils les ont récoltées (voir l’exemple du geai et des glands). On parle de zoochorie ou transport par des animaux. Pendant l’hiver, ils retrouvent ces réserves cachées pour les consommer mais en oublient toujours au moins quelques unes qui, au printemps suivant, en bonne position en plus, pourront germer avec plus de chances de se trouver dans un site qui permettra leur développement. Ainsi, de nouveaux individus peuvent s’installer et assurer le renouvellement de la population et son expansion éventuelle.

Jeunes plantules installées sur du bois mort : peut-être que les faînes y avaient été cachées par un oiseau ? 
Jeunes hêtres en lisière d’une hêtraie 
La relève est assurée avec ces jeunes d’au moins 2 ou 3 ans
Si une graine transportée depuis une population isolée atteint un autre population isolée et réussit à donner un nouvel individu, et si ce dernier réussit à se reproduire à son tour, alors on aura eu un événement de dispersion efficace qui pourra permettre un brassage génétique entre les deux populations (celle d’origine et celle colonisée). Cette probabilité dépend de nombreux facteurs : le nombre et l’efficacité des agents transporteurs (leur rayon d’action) ; la distance entre les populations isolées, la nature de la matrice paysagère qui les sépare (hostile aux transporteurs ?) ; présence de corridors favorables reliant les fragments ; … Souvent de tels évènements de dispersion à longue distance restent exceptionnels mais n’en sont pas moins cruciaux d’un point de vue génétique.

Etude de cas

Dans le nord-est de l’Espagne, en Catalogne, se trouve le parc naturel de Montseny, une région montagneuse dont les sommets au-dessus de 1000m sont peuplés de forêts dominées par le hêtre.
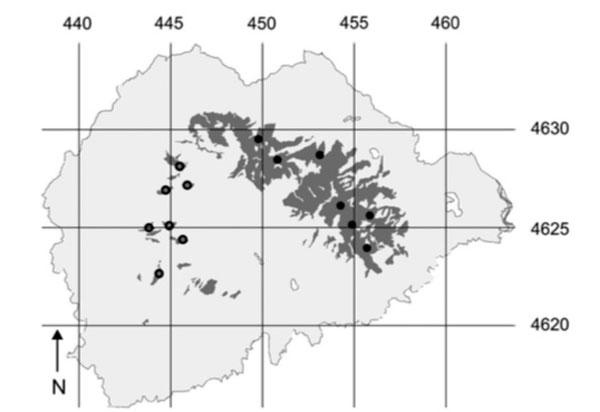
Gris clair = parc de Montseny ; noir = hêtraies ; cercles noirs = localisation des 7 échantillons prélevés dans des fragments ; points noirs : échantillons prélevés dans le massif continu.
Sur une chaîne au nord-est du parc qui culmine à plus de 1700m s’étend ainsi un vaste ensemble forestier de 2830hectares de forêts de hêtres presque continues. Vers le sud-ouest, sur une autre chaîne, un vaste secteur a été largement déforesté au 15ème siècle comme dans une majeure partie de l’Europe où l’on a un peu vite oublié que là aussi des surfaces considérables de forêts avaient été détruites à un rythme très rapide, du même ordre que celui que connaissent actuellement les forêts tropicales ! Une partie de notre développement économique a reposé sur cette destruction du manteau forestier originel. Dans cette chaîne plus basse, le hêtre se trouve confiné désormais dans une série de petits ilots forestiers que nous nommerons « fragments », entourés de pâtures et dominant une large plaine centrale cultivée : cet ensemble de fragments couvre au total 52 hectares et a donc au moins 600 ans d’âge. Belle occasion donc de comparer les hêtres des boisements restés continus et ceux des fragments isolés les uns des autres et à une certaine distance du massif continu.

Tout chercheur, avant de se lancer dans une étude, pose des hypothèses en fonction notamment de ce que l’on connaît déjà sur l’espèce étudiée ou à partir d’autres études du même type. On se dit donc ici que la fragmentation de l’habitat forestier doit provoquer des ruptures dans le système de reproduction et de dispersion du hêtre avec plusieurs conséquences possibles : une consanguinité croissante qui va donner une descendance de moins en moins viable, une forte différenciation génétique des petites populations isolées (chacune évoluant un peu plus ou moins en vase clos) et une érosion (ou baisse) de la variabilité génétique entre individus composant chaque population de fragment. A court terme, cela pourrait conduire à une baisse du succès reproductif dans les fragments et le risque d’extinction de ces populations ponctuelles ; à long terme, la réduction de la diversité génétique pourrait limiter les capacités d’adaptation face à la crise climatique en cours compte tenu des exigences écologiques assez strictes du hêtre.

Marqueurs
Pour savoir si les fragments échangent des individus entre eux (voir ci-dessus) ou avec le massif forestier continu lors de dispersions efficaces ou au contraire n’ont que très peu ou pas du tout d’échange (ne se reproduisant alors qu’entre individus de la même population), il faut faire appel aux outils dits moléculaires, i.e. à l’ADN qui conserve toutes les traces des flux de gènes via le processus de fécondation. Ils permettent d’estimer la diversité génétique des populations et de mettre en évidence les flux de gènes éventuels entre populations.

On utilise plus précisément des marqueurs appelés microsatellites : ce sont des gènes dispersés sur la chaîne d’ADN sur les chromosomes dans le noyau des cellules, formés de quelques centaines de paires de bases et n’ayant pas de fonction génétique connue (non codants). De ce fait, ils ne subissent pas de pression de sélection et sont en général très variables (nombreuses mutations) avec de nombreuses variantes ou allèles pour chacun d’eux. Si on sélectionne un certain nombre de ces microsatellites (6 dans le cas de cette étude sur le hêtre), chaque individu aura une combinaison propre d’allèles par rapport à ces 6 satellites ; ici, chaque gène possédait entre 3 et 12 allèles différents. Chaque individu (chaque hêtre donc), pour un même satellite ou gène, possède un allèle issu du « père » (ici, via le pollen des fleurs mâles) et un allèle issu de la « mère » (ici l’ovule du pistil de la fleur femelle). Si les deux allèles sont différents, on le qualifie d’hétérozygote ; s’ils sont identiques, il est homozygote (pour le gène considéré). Dans cette étude, on a donc prélevé 7 échantillons dans 7 fragments forestiers différents et 7 autres dispersés dans le massif continu : soit 14 échantillons dont l’ADN est extrait puis traité de manière à isoler les brins retenus. En combinant avec les 6 gènes étudiés, on a donc 84 combinaisons servant de points de comparaison !
Dans le cas d’une certaine consanguinité (reproduction entre individus de la même population), le nombre d’individus homozygotes pour un gène donné augmente : ainsi, on peut détecter si ce processus est installé et avec quelle intensité. Par ailleurs, à partir d’un modèle théorique d’une population où les reproductions se feraient au hasard (modèle de Hardy-Weinberg), on peut prédire les fréquences attendues des allèles et les comparer avec celles mesurées : tout écart traduira un « problème » sous-jacent comme la consanguinité par exemple.

Hêtres isolés : vestiges d’anciennes forêts continues en moyenne montagne 
Dans les pâturages, les vieux hêtres isolés témoignent souvent de défrichements très anciens
Résultats
Dans 6 cas (sur le total de 84) correspondant à des échantillons et des gènes différents, on a trouvé un net écart (voir ci-dessus) avec le modèle ; ils correspondent à des taux de consanguinité élevés et font tous partie des 7 échantillons prélevés dans des fragments forestiers isolés ; ce taux de consanguinité est deux fois plus élevé que dans les hêtres du massif continu, chez lesquels il n’est pas nul pour autant. Quand on compare les échantillons par paires dans le groupe fragments ou dans le groupe massif, le taux de différenciation génétique est 3 fois plus élevé pour les premiers que pour les seconds. Autrement dit, au sein du massif forestier, le flux de gènes maintient u niveau homogène et empêche la différenciation en sous-populations. Un test informatique montre de plus que les populations de cinq des fragments ont connu une période de forte réduction de leur abondance, passant par ce qu’on appelle un goulot d’étranglement ; aucun n’a été détecté dans les échantillons du massif continu. Plus les distances entre fragments forestiers sont grandes, plus les différences génétiques entre populations sont importantes ce qui n’est pas le cas pour le massif forestier bien que les échantillons aient été prélevés sur une très vaste surface.
Parmi les 102 allèles détectés sur l’ensemble des échantillons, près de 70% étaient rares ; mais, alors que 23 d’entre eux sont absents dans les échantillons des fragments, seulement 5 sont absents des échantillons du massif continu.

Les plantations de résineux fragmentent les boisements « de l’intérieur » 
Très ancienne clairière habitée qui a troué le manteau forestier originel
Diagnostic
Tous ces résultats convergent vers un constata évident : la fragmentation des forêts de hêtres, vieille de plus de 600 ans, a eu un effet significatif fort sur les populations isolées dans des fragments. A l’inverse, au sein du massif continu, le flux de gènes entre individus est resté fort : on parle de situation panmictique (mot dérivé de panmixie : reproduction au hasard).

Plus les fragments sont isolés et éloignés du massif continu, plus on assiste à une rupture de cet état panmictique vers une prédominance de la consanguinité. Ceci conduit à un statut de non-équilibre qui se traduit notamment par l’existence passée de goulots d’étranglement où certaines des populations ont peut être failli s’éteindre. Pourtant, ces fragments étudiés ont une taille de l’ordre de plusieurs dizaines d’hectares : cet effet résulte donc de leur isolement génétique et de leur passage par des phases de population très réduite ; ainsi, quand ils se sont reconstitués, les nouveaux arbres provenaient de quelques géniteurs apparentés en vase clos.
Lors des prélèvements, on a sélectionné dans deux des fragments des vieux arbres et des jeunes arbres : ces derniers ont une diversité génétique moindre et un taux de consanguinité plus élevé que les anciens. Cela veut dire qu’au moins dans ces deux fragments, la population ne s’est pas remise de son goulot d’étranglement et n’a pas « reçu de sang neuf ».
Le plus surprenant dans ces résultats, c’est que le hêtre pratique fortement l’auto-incompatibilité (voir second paragraphe) et la pollinisation par le vent permet a priori des transports à longue distance ; or, les taux de consanguinité observés ici sont bien plus élevés que ceux par exemple trouvés dans une étude sur des chênes sessiles en Irlande. Ici, les populations des fragments se sont nettement plus différenciées. Sans doute que l’ampleur de la déforestation passée a conduit à un très fort isolement des fragments : l’apport de gènes depuis les massifs continus encore en place ne se fait plus et ce, même dans des fragments de plusieurs hectares.
Mal-hêtre ?
Il reste une question centrale : est-ce grave ? Cette perte de diversité génétique associée à la consanguinité croissante peut avoir des effets directs notamment sur la viabilité des graines par accumulation de mutations négatives chez les individus homozygotes : des taux de germination moindres, une forte mortalité des plantules, … Et puis, face à la crise climatique en cours, dont les effets sont encore plus accentués sur les fragments (surface de contact plus grande au niveau des lisières), le potentiel de sélection de nouveaux individus avec des caractères susceptibles de les rendre plus résistants aux nouvelles conditions va se trouver réduit par manque de diversité. Autrement dit, ces populations fragmentées ont bien plus de risques de disparaître assez rapidement par rapport à celles des massifs compacts, même si cela se fera avec un certain décalage dans le temps compte tenu de la longévité des hêtres (plusieurs centaines d’années). Les politiques de conservation de l’environnement devront prendre prioritairement en compte ces zones fragmentées, notamment en récréant quand c’est possible des corridors boisés reliant ces fragments entre eux et avec le massif continu le plus proche quand c’est possible.

Bibliographie
Genetic effects of chronic habitat fragmentation in a wind-pollinated tree. Alistair S. Jump and Josep Penuelas 8096–8100 ? PNAS ; May 23, 2006 ; vol. 103 ; no. 21.
Structuration génétique des populations de tritons palmés – A. Trochet. Université de Toulouse – 2014/2015. Association BUFO : excellent document pédagogique sur la méthode d’étude de la structure génétique des populations.
A retrouver dans nos ouvrages
Page(s) : 188-189

