Le févier d’Amérique : une dispersion anachronique ?
Gleditsia triacanthos
La notion d’anachronisme de dispersion est née dans les années 1980 à partir d’observations sur des fruits charnus dans les forêts d’Amérique centrale : certains d’entre eux réunissent des caractéristiques (des traits) associées à une dispersion via de très grands mammifères capables de les avaler tout entiers, de les digérer puis de rejeter leurs graines intactes dans les excréments : on parle de syndrome de dispersion par la mégafaune (mammifères de plus de une tonne) (voir la chronique sur ce thème). Sauf que les dits mammifères, dans cet environnement là, n’existent plus : cette mégafaune, très diversifiée, a disparu entièrement en Amérique du sud et centrale il y a un peu plus de 10 000 ans (voir la chronique sur cette mégafaune). Les traits ainsi conservés par les fruits apparaissent alors comme des « fantômes » de l’existence passée de ces géants disparus.

En Amérique du nord aussi, il y a eu aussi un tel processus d’extinction en masse de la mégafaune (voir en bas de chronique dans Ouvrages le dossier sur ce thème) et on y suspecte plusieurs espèces d’arbres produisant des fruits charnus de relever de ce syndrome, dont le févier d’Amérique, un arbre ornemental de plus en plus cultivé en ville et bien connu pour ses gousses géantes contenant des dizaines de grosses graines et une pulpe verte sucrée (voir la chronique sur la reproduction du févier). Nous allons parcourir les arguments pour et contre cette hypothèse car il subsiste encore un vif débat quant à sa validité.
Une dispersion actuelle un peu « faiblarde »
Déjà, au niveau de la production de graines, le févier se distingue par son caractère dioïque (sexes séparés) et, pour une population donnée, une production annuelle qui dépend de quelques individus femelles, différents d’une année à l’autre (voir la chronique sur la reproduction du févier). Le grand spécialiste de l’espèce, A. Schnabel, a montré que dans deux populations du Kansas (1), séparées mais dans un même contexte écologique, la dispersion était pourtant très différente : dans l’une, on observait une descendance (des plantules) très agglomérée au pied des arbres mères, résultat d’une simple chute des grands fruits au sol ; dans l’autre population, au contraire, la descendance se trouvait plus ou moins éloignée des arbres mères, du fait du passage de cerfs de Virginie qui avaient consommé les gousses au sol. Donc, la dispersion semble très aléatoire.
Dans une autre étude sur une troisième population, le même auteur (2) montre que celle-ci est structurée à une échelle fine selon des groupes d’individus très rapprochés et en plus très apparentés, témoins d’une dispersion à très courte distance : les fruits sont peu déplacés par rapport aux arbres mères, au plus jusqu’à l’arbre le plus proche ! Tout ceci contraste fortement avec « l’investissement » énergétique important dans la production de très gros fruits nombreux, chargés en matières nutritives et en graines … tout ceci pour un si piètre résultat ?
-

- Gousses de févier tombées au pied de l’arbre mère par simple gravité.

Un autre indice d’une dispersion naturelle peu efficace peut se lire dans la répartition originelle du févier : à l’arrivée des colons européens, il se trouvait localisé dans le centre-est des U.S.A., le long de vallées alluviales, comme si après les dernières glaciations qui avaient repoussé nombre d’espèces plus au sud, il avait été incapable de reconquérir de nouveaux territoires vers le nord comme l’ont fait nombre d’autres espèces. Ce confinement en milieu alluvial paraît d’autant plus étrange que depuis son utilisation comme arbre ornemental, on s’est rendu compte qu’il supportait très bien la sécheresse ! A priori, il n’est donc pas limité par un problème d’humidité du sol.
Par contre, depuis la colonisation humaine, le févier a connu une formidable expansion géographique, à partir des arbres plantés en ville ou comme brise-vents dans les plaines ou comme fourrage (par le feuillage) y compris hors de son pays d’origine : en Australie, il est devenu très invasif et indésirable tout comme au Chili, en Argentine (3) ou en Afrique du sud. Donc, ce n’est pas une faiblesse dans la reproduction elle-même qui limitait à l’origine le février. On voit donc d’emblée une série d’anomalies relatives dans la capacité de dispersion du févier.
-

- Févier d’Amérique planté dans un parc

-

- Troncs de Févier dans un parc : de loin, l’écorce grise se reconnaît à ses « crêtes » dressées

Une convergence d’indices
C. Barlow a consacré un ouvrage entier, passionnant, à ce problème des anachronismes de dispersion sous le titre « The ghosts of evolution » (voir Bibliographie : 4) où il a élaboré un véritable plaidoyer en faveur de l’idée de fruits anachroniques chez le févier, entre autres. Voyons sa liste d’arguments :
– la taille des fruits (jusqu’à 50cm de long) : des mégafruits ! Pourquoi des fruits aussi volumineux pour une douzaine de graines ? Pour une dispersion efficace, la gousse doit être mangée d’un seul bloc : si elle est fragmentée en morceaux, il y aura des pertes de graines au cours du processus ; il n’existe plus aucun animal vivant en Amérique du nord capable d’avaler de tels fruits
– des fruits qui ne s’ouvrent pas (indéhiscents) : les graines restent hors d’atteinte même sur des fruits tombés au sol ce qui limite la prédation ; il faut manger le fruit pour accéder aux graines
-

- Gousses en automne

-

- En hiver, une partie des gousses persiste longtemps sur l’arbre dénudé

– la coloration brun pourpre des gousses
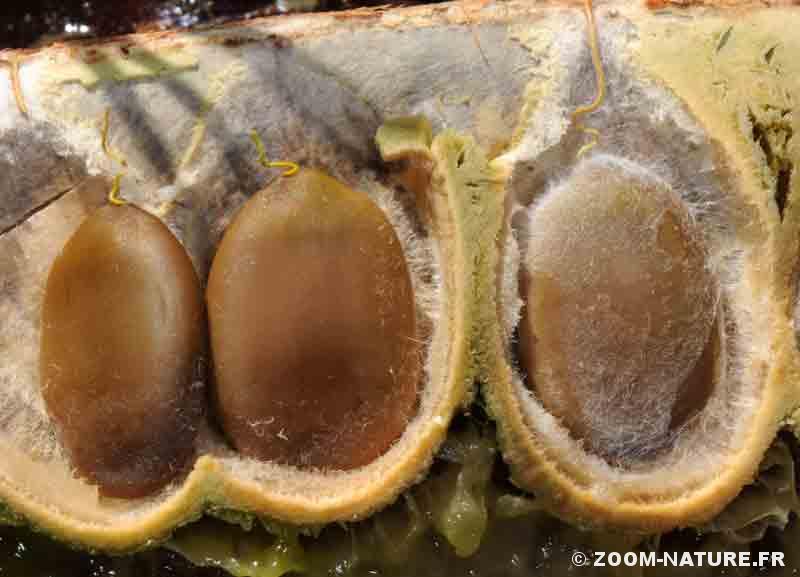
– la dormance profonde des graines liée à un tégument très épais et résistant ; en culture, on conseille de les tremper une à deux heures dans de l’eau très chaude avec de …. l’acide sulfurique pour espérer les voir germer ensuite assez rapidement ; certes ce caractère se retrouve chez de nombreuses autres légumineuses, mais, ici, il est porté à un paroxysme ; ceci laisse donc à penser que ce revêtement protège les graines contre les dents broyeuses et le suc digestif ; il semble même que le passage « à travers un animal » lève en partie cette dormance profonde en altérant le tégument
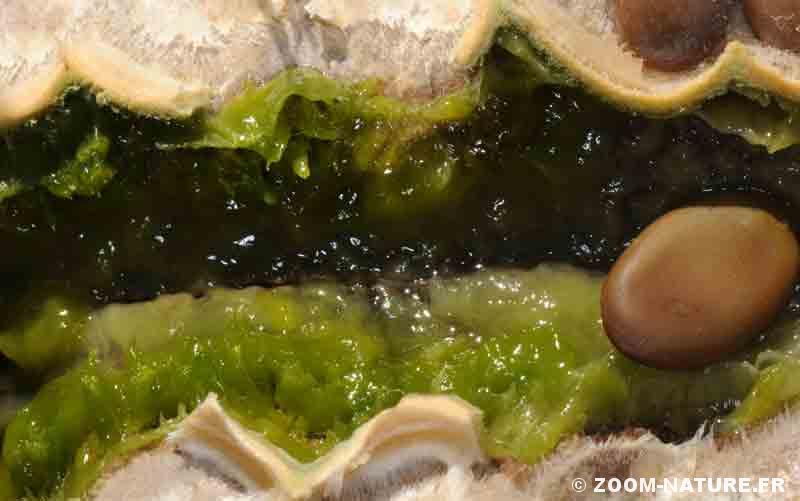
– la pulpe sucrée et riche en protéines contenue dans la gousse, disponible pendant tout l’hiver puisqu’une partie des gousses reste sur l’arbre, accessible à de grands animaux terrestres
-

- Gousse géante de févier (jusqu’à 50cm de long) avec sa pulpe et ses graines
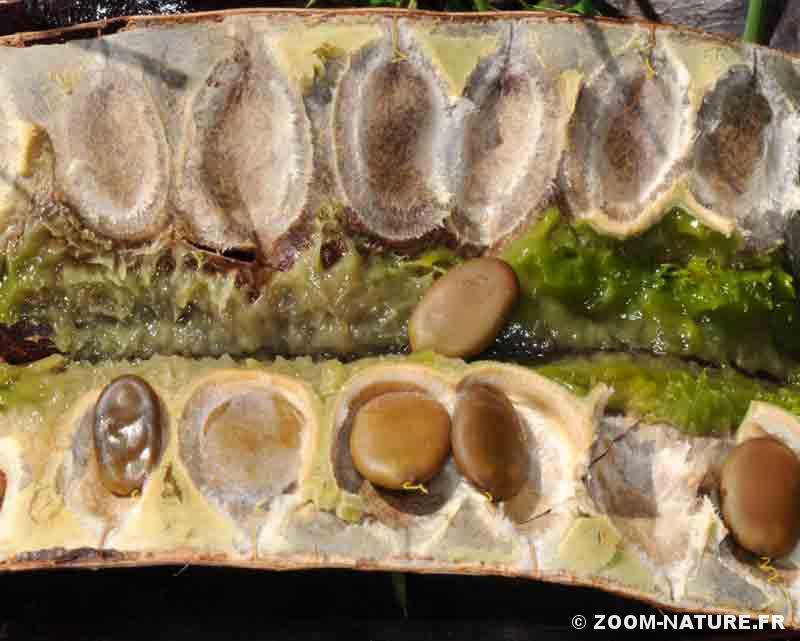
-

- Pulpe verte sucrée à odeur fermentée.
-

- Graines plates à tégument cireux et très épais et résistant
– les fortes épines ramifiées qui hérissent le tronc des jeunes arbres jusqu’à cinq mètres de hauteur alors qu’en Amérique du nord, il n’y a pas d’animaux arboricoles susceptibles de grimper dans l’arbre ; de telles épines (que l’on retrouve d’ailleurs chez divers arbres ou arbustes américains) serviraient à empêcher les grands herbivores attirés par les fruits d’en profiter pour manger aussi le feuillage très nutritif (souvent utilisé comme fourrage) ou de s’attaquer à l’écorce comme le font couramment les chevaux.
-

- Epines ramifiées très puissantes sur le tronc, dans la partie inférieure
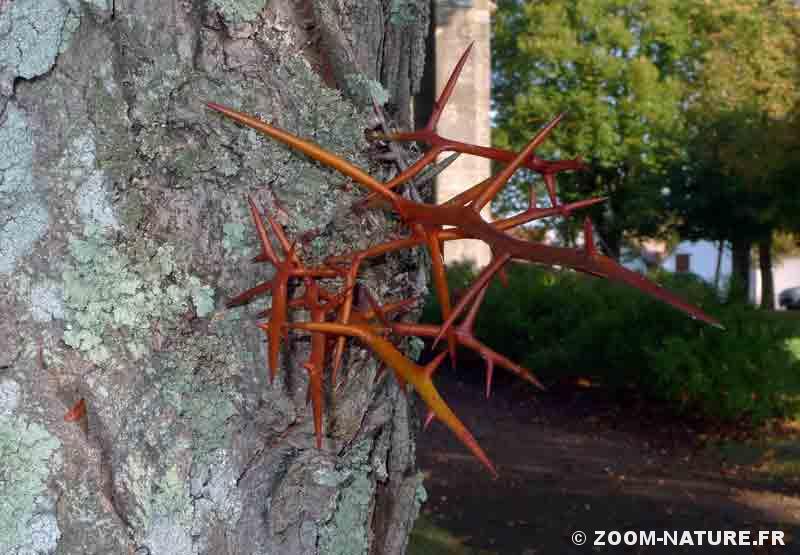
-

- Bouquets d’épines répartis tout autour du tronc

-

- Le manchon d’épines localisé dans les cinq premiers mètres représente une barrière dissuasive à toute approche !

Tous ces traits convergent donc vers le syndrome de dispersion par la mégafaune disparue. Mais, on peut avancer des objections et notamment le fait qu’aucun de ces traits n’est une preuve absolue en soit : la dureté des graines peut par exemple être interprétée aussi comme une protection contre la dent des rongeurs qui sont des prédateurs ; la grande taille des fruits tordus et plats peut aussi être un moyen de dispersion par le vent une fois tombés au sol, le vent pouvant les faire rouler sur la neige, …..
La dispersion secondaire
Les objections renvoient aux autres possibilités de dispersion car, même actuellement, le févier réussit à être dispersé de plusieurs manières. La flottaison (permise entre autres par la fermeture hermétique des fruits et leur paroi coriace) reste un moyen de dispersion efficace mais uniquement le long des cours d’eau, ce qui explique d’ailleurs la localisation originelle de cette espèce. La simple chute au sol constitue en soi un mode de dispersion mais elle conduit à une agrégation des descendants sous l’ombre des parents, peu efficace génétiquement, ce qui est effectivement observé. Des animaux actuels de taille moyenne tels que les cerfs de Virginie mais aussi surtout le bétail et les chevaux consomment volontiers ces fruits et on retrouve les graines dures dispersées dans leurs excréments ; mais, beaucoup de graines sont recrachées sur place au cours du repas ou tombent au sol sur place. On peut donc conclure qu’en fait, le févier présente sans aucun doute un anachronisme partiel de dispersion qu’il réussit à compenser plus ou moins par des moyens de dispersion secondaire et, surtout, par l’intervention humaine qui a propagé très activement cet arbre et a permis son expansion récente, y compris hors de son continent natal.
Pour comprendre cette relation févier/mégafaune, il faut se replacer dans le contexte évolutif et donc interroger l’histoire du genre et les contextes faunistiques et écologiques dans lesquels a eu lieu cette évolution : ceci sera l’objet d’une troisième chronique sur l’évolution des féviers.
BIBLIOGRAPHIE
- Understanding the population genetic structure of Gleditsia triacanthos L.: seed dispersal and variation in female reproductive success. A. SCHNABEL, J. D. NASON, and J. L. HAMRICK. Molecular Ecology (1998) 7, 819–832
- Comparative genetic structure of two co-occurring tree species, Maclura pomifera (Moraceae) and Gleditsia triacanthos (Leguminosae). A. Schnabel ; R.H. Laushman ; J.L. Hamrick. Heredity 67 (1991) 357-364
- Establishment of Honey Locust (Gleditsia triacanthos) in Burned Pampean Grasslands. E. J. CHANETON, C. NOEMI MAZIA, M. MACHERA, A. UCHITEL, and C. M. GHERSA Weed Technology. 2004. Volume 18:1325–1329
- The ghosts of evolution. Non sensical fruit, missing partners, and other ecological anchronisms. C. Barlow. Basic Books. 2000.
A retrouver dans nos ouvrages
Page(s) : 444-454

Page(s) : 144-145

