Le butome en ombelle ou jonc fleuri
Butomus umbellatus

22.12.2020 Le jonc fleuri (nom populaire) ou butome en ombelle (nom officiel) est une plante du bord des eaux douces, répandue mais peu commune et en régression dans notre pays. Elle mérite le détour avec sa floraison superbe d’un beau rose soutenu et ses inflorescences en ombelles très fournies. Cette plante a fait l’objet d’une foule d’études quant à ses modes de reproduction car elle a été introduite en Amérique du nord où elle est devenue très invasive et problématique. Entrons donc dans l’intimité de cette belle du bord des eaux.


Ombelles

Touffe fleurie au milieu de la végétation aquatique 
Touffes de feuilles
Le butome forme le plus souvent des touffes séparées parfois en colonies très denses ; elles se composent de nombreuses feuilles longues (jusqu’à 1m), d’un vert franc, plus ou moins disposées sur deux rangs (distiques) comme chez les iris et triangulaires en section (triquètres) comme celles des rubaniers ou des laîches. Elles s’élargissent progressivement vers le haut et tendent à se torsader vers leur extrémité. A ce stade, le butome peut se confondre avec une foule d’autres plantes aquatiques si on n’a pas un œil exercé. Par contre, dès la période de floraison, au cœur de l’été, l’identifier devient un jeu d’enfant et on peut le reconnaître même en roulant en voiture tellement sa floraison est unique avec une apparence presque exotique.

Ombelles à nombreux rayons 
Trois bractées foncées à la base de l’ombelle
De la touffe de feuilles (voir ci-dessous) émergent de longues tiges florales nues cylindriques et robustes (jusqu’à 1,50m) ; elles sont remplies à l’intérieur d’espaces aérifères, ce qui leur donne un aspect interne spongieux, dispositif classique chez de nombreuses plantes aquatiques, et elles renferment de nombreuses micro-baguettes cristallines. Chaque tige florale (hampe ou scape) se termine par une ombelle très étalée de 15 à 30 fleurs, voire jusqu’à 50, d’un beau rose soutenu. Chaque rayon de l’ombelle correspond au long pédoncule d’une fleur ; il s’agit donc d’une ombelle simple. Trois petites feuilles (bractées) sous-tendent la base de l’ensemble des rayons comme un involucre. Si le vocabulaire utilisé ici rappelle celui concernant les ombellifères ou apiacées (voir la chronique sur cette famille), cette plante n’a strictement rien à voir avec cette famille (voir ci-dessous). D’ailleurs, au moment de l’initiation de cette ombelle, à l’aisselle de chacune des trois bractées basales se forme un massif cellulaire indifférencié (méristème) qui engendre sa propre partie de l’ombelle qui a donc une origine triple inhabituelle.
Multiples de 3

Le chiffre 3 évoqué ci-dessus se retrouve dans tous les détails de la structure de ces belles fleurs d’un diamètre de 2 à 3cm. En partant de l’extérieur, on a deux cercles successifs de trois pièces florales (le périanthe) d’un très beau rose mais sensiblement différentes : le cercle externe qui correspond au calice aux trois pièces plus courtes parfois teintées de verdâtre à leur extrémité puis le cercle interne, équivalent de la corolle de trois pièces d’un rose plus vif et veinées de foncé. Cette structure est typique du groupe des Monocotylédones dans lequel s’insère la famille du butome (voir ci-dessous) et en fait la distinction calice/corolle n’a plus guère de sens : on parle de tépales pour désigner ces six pièces ressemblant à des pétales (pétaloïdes).
Viennent ensuite les neuf étamines disposées en deux cercles (verticilles) successifs : un de six puis un de trois. Portées sur des filaments aplatis, les anthères frappent par leur couleur rouge foncé qui contraste sur le fond rose de la fleur. Elles s’ouvrent par des fentes latérales et libèrent un pollen jaune avant de faner.

Enfin, tout au centre, viennent les pistils au nombre de six en un seul cercle : le gynécée ; ils sont libres mais serrés, adhérant à leurs bases par des poils entremêlés. Chaque pistil comporte un ovaire à une loge renfermant de nombreux ovules, un style un peu arqué qui se prolonge en un stigmate devenant jaune quand il est réceptif au pollen et ressemblant alors aux anthères ouvertes ! A la base des pistils se trouvent des glandes (nectaires) qui produisent un abondant nectar libéré pendant toute la floraison.

Les pistils fécondés deviennent des fruits secs avec une seule ligne d’ouverture : on parle de follicules comme chez, par exemple, les hellébores (voir la chronique). A maturité, ils s’ouvrent par la fente ventrale et libèrent leurs nombreuses graines.
Pollinisation

Les fleurs du butome présentent tous les caractères d’une plante entomophile, i.e. pollinisée par des insectes : la forme de coupe des fleurs, le nectar abondant et le pollen faciles d’accès dans des inflorescences denses et fournies en plates formes étalées les rendent accessibles à des espèces d’insectes généralistes non spécialisées (pas besoin d’une trompe ou d’une langue longue pour accéder au nectar). Effectivement, il semble que les visiteurs principaux soient des mouches dont des syrphes (voir la chronique sur ces pollinisateurs méconnus) et des abeilles domestiques ou solitaires.

Mouche butinant sur une fleur de butome ; noter les stigmates mûrs bien jaunes 
Grisette (Hespéride) visitant des fleurs de butome
On a récemment documenté dans le nord-est de la Chine l’existence de populations avec un gynécée central blanc au lieu du rose vif classique ; une étude de suivi des pollinisateurs et de la production de graines montre que ces fleurs à cœur blanc n’attirent pas plus les visiteurs que celles à cœur rose ni des visiteurs différents ; la forme blanche semble en fait liée au niveau de l’eau dans le milieu de vie.

Au niveau individuel, chaque fleur passe par trois stades de un jour chacun en moyenne. Le premier jour, les anthères s’ouvrent et libèrent le pollen alors que les stigmates des pistils ne sont pas du tout réceptifs ; on parle de phase mâle. Le second jour, il n’y a plus de pollen disponible mais les stigmates restent inactifs ; on parle de phase neutre. Enfin, le troisième jour, les stigmates virent au jaune et deviennent réceptifs : c’est la phase femelle. La fleur est donc « mâle » d’abord est qualifiée de protandre (andros = masculin). L’existence d’une phase neutre intermédiaire est originale et rare car chez beaucoup de plantes protandres, on a plutôt un passage progressif avec chevauchement partiel des deux phases mâle et femelle (voir l’exemple de la saponaire). Le dépôt de pollen sur les stigmates précipite la fin de la phase femelle finale.

Fleur juste éclose au stade mâle 
Fleur au stade mâle : les anthères sont ouvertes 
Fleur au stade neutre : anthères vides et stigmates déployés mais encore blancs (non réceptifs)
Au niveau collectif d’une ombelle, on constate de même une certaine synchronisation qui réduit les chevauchements des phases mâle et femelle entre fleurs de la même ombelle. En moyenne, plus de 70% des fleurs d’une ombelle finissent leur phase femelle alors qu’aucune autre fleur de l’ombelle ne se trouve en phase mâle (et donc susceptible d’apporter du pollen venant de la même plante) ; un suivi sur plusieurs populations confirme que 87% des ombelles sont au même stade soit mâle soit femelle. Ce trait a toute son importance quand on sait que les insectes pollinisateurs tendent à visiter successivement plusieurs fleurs proches d’une même ombelle avant de passer à une autre. Cette séparation temporelle des sexes (dichogamie) limite donc sérieusement les risques de fécondation en interne au sein d’une fleur ou entre fleurs de la même ombelle, ce qui présente des désavantages génétiques importants.
Clonale

Cependant, en pratique, quand on étudie la production effective de graines, on découvre une situation très variable : certains individus (touffes) ne produisent pas ou très peu de graines tout en ayant des fleurs fonctionnelles alors que d’autres en libèrent « beaucoup ». Sur le terrain, on observe qu’en fait le recrutement de nouveaux pieds se fait très peu à partir de germinations de graines et de nouvelles plantules. Les graines libérées peuvent flotter sur l’eau et être emportées mais souvent, en automne au moment de leur libération, les milieux de vie sont partiellement asséchés. La germination printanière ou estivale des graines ne se fait qu’après un séjour hivernal dans le sédiment et une immersion dans l’eau (inondation d’un milieu asséché en automne). Les jeunes plantes ont besoin d’un substrat mouillé et en plein soleil pendant au moins deux mois et leur croissance reste lente.
Toutes ces contraintes limitant la dispersion et le renouvellement par les graines n’empêchent pourtant pas cette plante d’être capable de coloniser activement certains milieux, voire même de les envahir (voir ci-dessous). Le butome présente en effet des dispositions pour la multiplication végétative, i.e. une forme de reproduction sans passer par la reproduction et les graines.
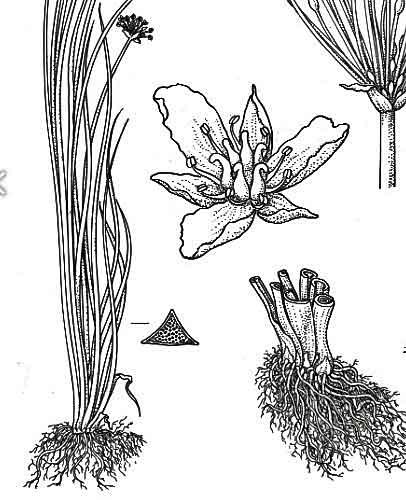
L’appareil souterrain se compose d’un rhizome (tige souterraine) assez épais qui s’étend progressivement et donne de nouvelles plantes ; mais surtout, sur ce rhizome, se forment des structures en forme de bulbilles, des bourgeons latéraux attachés de manière très lâche. A la moindre perturbation du sédiment meuble (vase) qui enveloppe les rhizomes, ces bulbilles peuvent se détacher et être emportés par le courant ou déplacés et peuvent, plus loin, redonner une nouvelle plante chacun. Ainsi se créent des peuplements étalés clonaux, i.e. identiques génétiquement aux parents donneurs de bulbilles. Les perturbations favorables à ce processus peuvent être variées : une crue de rivière qui érode le fond vaseux ; un cygne ou des oies ou un ragondin qui fouille la vase pour prélever des fragments ; le curage d’un fossé ou le passage d’un engin à moteur ; … Ajoutons à tout cela, un autre mode de multiplication récent : la culture par l’homme du butome comme espèce ornementale souvent plantée près de cours d’eau ou de plans d’eau et qui ensuite s’échappent très facilement !
Dédoublement
Mais la réalité est encore plus complexe car à ce double jeu reproduction sexuée par graines/multiplication végétative vient se superposer une étonnante particularité génétique de cette espèce. On a découvert qu’il existait en fait deux types de populations semblables morphologiquement mais différentes par le nombre de chromosomes contenu dans chacune de leurs cellules (caryotype). Les unes ont 13 paires (2n = 26) de chromosomes différents (comme nous espèce humaine en avons 23 paires dont la paire de chromosomes sexuels XX ou XY) et sont qualifiés de diploïdes (diplo = deux) ; les autres ont 13 triplets (3n = 39) des mêmes chromosomes et sont dits triploïdes. Des études minutieuses menées notamment en Europe centrale ont permis de déceler de nombreuses différences dans l’écologie et le développement de ces deux types de populations, résumées dans le tableau ci-dessous.

Outre la production de bulbilles dans les inflorescences, à la manière de nombreux ails sauvages, une autre forme de multiplication végétative, les triploïdes se démarquent par leur système reproducteur : leurs fleurs sont auto-incompatibles, i.e. que la fécondation ne peut se faire qu’entre ovules d’une plante et pollen venant d’une autre plante ; or, comme les triploïdes forment souvent des peuplements clonaux, cette reproduction croisée devient quasi impossible. Si on les pollinise manuellement avec du pollen venant d’un autre pied triploïde, on réussit à obtenir des graines : la barrière est donc interne. Les individus diploïdes sont par contre autocompatibles : en cas d’absence d’autre pied dans le voisinage, ils peuvent quand même produire des graines.
En Europe, il semble que la forme triploïde domine ce qui explique la faible régénération par graines constatée et les populations semblent uniquement formées d’un seul type (pas de mélange) à la fois. Les triploïdes allouent l’essentiel de leurs ressources nutritives à produire des rhizomes puissants et des bulbilles mais ne produisent pas ou très peu de graines (tout en conservant des fleurs viables !). Les diploïdes, eux, investissent leurs ressources pour un quart dans les inflorescences et la production de graines mais aussi à 40% dans la fabrication de bulbilles en optant pour des rhizomes et appareils végétatifs moins robustes ; ils ont donc un double avantage à la capacité de propagation alors que les triploïdes se maintiennent plus longtemps et s’étendent lentement.
Invasive
L’aire d’origine du butome couvre l’Eurasie tempérée jusqu’en Scandinavie au nord. En Amérique du nord, il a été introduit comme ornementale à la fin du 19ème siècle et n’a pas tardé à s’échapper et à se naturaliser surtout dans la région des Grands Lacs et le bassin du Saint-Laurent. Assez rapidement, il a pris un caractère invasif agressif colonisant les bords des lacs en peuplements exclusifs denses et les canaux d’irrigation. Sa présence concurrence sérieusement les espèces autochtones et pose de nombreux problèmes humains : circulation de l’eau dans les canaux, envasement, impact sur la faune aquatique, gêne pour la navigation, … L’invasion résulte clairement d’introductions répétées en de nombreux endroits. Mais au fur et à mesure que l’espèce s’est installée, on a découvert que les triploïdes étaient rares (contrairement à ce qui se passe en Europe) et que la majorité des populations était diploïde, i.e. produisant facilement des graines. Un pied de butome peut produire jusqu’à 20 000 graines par an, plus les bulbilles des rhizomes. Comme il a trouvé des habitats favorables (eaux peu profondes), il a donc pu se propager très rapidement. On suppose que les plants ornementaux sélectionnés pour la qualité de leur floraison sont majoritairement diploïdes ce qui a favorisé leur expansion au départ. On peut imaginer qu’en Europe, via l’augmentation des introductions artificielles, les populations diploïdes ne se répandent plus mais elles semblent imitées par la disponibilité en milieux favorables dont la qualité se dégrade.
Aquatique

Le butome colonise les milieux aquatiques avec une couche de vase propice à son installation. On le trouve souvent au sein de zones marécageuses étendues dans des petits plans d’eau stagnante avec des niveaux d’eau fluctuant fortement chaque année. Ainsi, il prospère dans les fossés et canaux peu profonds à condition qu’ils ne soient pas curés trop souvent et trop profondément ce qui élimine les rhizomes. Il pousse le plus souvent les pieds dans l’eau avec les feuilles en partie émergées. On le retrouve dans les petites rivières et canaux importants où il peut vivre entièrement submergé (jusqu’à 3m de profondeur) sous une forme très modifiée, forcément stérile sous l’eau, avec de longues feuilles souples rubanées ; ceci s’observe chez de nombreuses autres plantes aquatiques émergées comme la sagittaire par exemple ou les nénuphars.

Mare temporaire presque complètement occupée par les butomes 
Il côtoie souvent les iris d’eau (en arrière-plan)
Il semble favorisé par une baisse du niveau d’eau en été car le sédiment se réchaufferait mieux et augmenterait le développement des rhizomes. Si le niveau d’eau ne baisse qu’un peu, des ceintures se forment le long de la frange partiellement asséchée en été. Inversement, une hausse prolongée du niveau d’eau freine son développement même s’il réussit à survivre sous forme végétative.
Le butome préfère les sols alluviaux ou argileux surtout dans les régions calcaires (notamment les populations triploïdes) et ne monte pas en altitude. En France, on le trouve un peu partout mais en populations très dispersées et en régression ; au sud d’une ligne Bordeaux/Lyon (moitié sud) il se cantonne plus en milieu fluviatile avec ces formes submergées plus difficiles à repérer du fait de leur ressemblance avec diverses autres plantes aquatiques.
A noter qu’en Asie centrale, on consomme les rhizomes riches en amidon sous forme de farine ou cuit en pains ou galettes. On peut aussi torréfier ces rhizomes pour fabriquer un ersatz de café, pratique répandue avec diverses autres plantes aquatiques à rhizomes dont les massettes.
Jonc, vraiment ?

En dehors de l’adjectif fleuri qui lui va bien, le surnom populaire de jonc apparaît bien malencontreux à double titre. D’une part, aucun jonc n’a de feuilles triangulaires en section ni de telles fleurs aussi développées ; d’autre part, la famille des joncs (Joncacées) n’est pas du tout apparentée à celle du butome (Butomacées). Les joncacées s’insèrent dans la lignée des Poales avec les Graminées et les Cypéracées (laîches, linaigrettes, scirpes, souchets, …) aux fleurs « écailleuses ». La famille des butomacées, qui ne renferme que cette seule espèce, s’inscrit quant à elle dans une toute autre lignée, les alismatales, un ensemble hétérogène de plantes toutes aquatiques dont les seules plantes vertes marines comme les zostéracées.

Quatre Hydrocharitacées : la morène petit-nénuphar 
La petite naïade 
Le lagarosiphon élevé 
L’élodée du Canada
Dans cette lignée, les butomacées sont étroitement apparentées à la famille des hydrocharitacées qui compte dans ses rangs les élodées, le stratiote faux-aloès, les vallisnéries ou les naïades.

Elles forment avec la famille des Hydrocharitacées un groupe lui-même apparenté aux Alismacées (plantains d’eau, sagittaires, …). Le butome (et sa famille) se démarque nettement par ses fleurs très développées et ses fruits formés de six follicules réunis en couronne.

Deux alismacées : la sagittaire de Montevideo 
Le plantain d’eau
Le nom étrange de butome vient de deux racines : bous pour bœuf et temnos= je coupe. Elles font allusion à des plantes aquatiques aux feuilles triquètres comme celles du butome mais très coupantes au toucher car enduites de silice : les laîches ou carex (voir la chronique sur les « rouchus« ) ; d’ailleurs, à tout prendre, il aurait mieux valu surnommer le butome laîche fleurie ! Quant au bœuf, outre le fait que les bovins peuvent brouter ces plantes, on peut aussi le relier à un autre nom d’un oiseau des marais, un héron nocturne, le butor étoilé : son chant ressemble à un mugissement profond et lui a valu le surnom de bœuf des marais. D’où le rapprochement phonétique de ces deux noms : de quoi composer une fable intitulée Le butor et le butome !

Bibliographie
Aquatic plants in Britain and Ireland. C.D. Preston and JM Croft. Harley Books 2001
Contrasting patterns of spatial genetic structure of diploid and triploid populations of the clonal aquatic species Butomus umbellatus (Butoaceae) in Central Europe. Jan Kirschner et al. Folia Geobotanica 39: 13–26, 2004
The biology of Butomus umbellatus in shallow waters with fluctuating water level. Z. Hroudovda et al. Hydrobiologia 340: 27-30, 1996.
Germination responses of diploid Butomus umbellatus to light, temperature and flooding. Zdenka Hroudová et al. Flora (2003) 198, 37–44
Fluctuating selection by water level on gynoecium colour polymorphism in an aquatic plant. Xiao-Xin Tang and Shuang-Quan Huang Annals of Botany 106: 843–848, 2010
Functionnal analysis of synchronous dichogamy in flowering rush Butomus umbellatus (Butomaceae). MICHAEL BHARDWAJ AND CHRISTOPHER G. ECKERT. American Journal of Botany 88(12): 2204–2213. 2001
Evolutionnary increase in sexual and clonal reproductive capacity during biological invasion in an aquatic plant Butomus umbellatus (Butomaceae) JEREMY S. BROWN AND CHRISTOPHER G. ECKERT American Journal of Botany 92(3): 495–502. 2005.
The complete chloroplast genome of Butomus umbellatus L. and its phylogenetic position. Zhaoping Yang & Luxian Liu (2019), Mitochondrial DNA Part B 4:2, 3700-3701,
