Les nids d’oiseaux de la carotte sauvage
Daucus carota subsp.carota var. carota
29/08/2022 Nous consacrons quatre chroniques à la carotte sauvage, histoire de démontrer que même une espèce ultra-banale peut être un sujet d’intérêt par différents aspects de sa biologie. Ainsi, nous avons d’abord présenté le cycle de vie bisannuel dans une première chronique ; puis, nous avons zoomé sur la floraison et la pollinisation dans une seconde chronique. Ici, nous allons poursuivre en nous intéressant au devenir des ombelles aux fleurs fécondées, i.e. la fructification et la libération des graines et leur dispersion. La carotte sauvage s’y singularise par une évolution bien particulière de ses ombelles fructifiées ; la dispersion finale des graines apporte aussi son lot d’originalités.

Diakènes

Au moment de la floraison, les ombelles adoptent majoritairement un port très aplati qui en fait des plates-formes d’atterrissage pour pollinisateurs de toutes sortes (voir la chronique Corne d’abondance) ; ce port est permis par les rayons qui sous-tendent chacun une ombellule de fleurs et s’orientent selon un angle propre et s’allongent plus ou moins, selon leur position au sein de l’ombelle. En général, seule l’ombelle terminale et les ombelles latérales d’ordre 1 (voir la chronique) sur une tige donnée atteignent le stade de fructification ; comme il y a un décalage d’environ deux semaines dans la floraison successive des différents ordres d’ombelles, celles de second ordre et au-delà fructifient rarement.

Pour une ombelle donnée, toutes les fleurs éclosent simultanément ce qui renforce l’effet attractif de « super-fleur ». Dès la fin de leur floraison, alors que les pétales ne sont pas encore tous tombés, on note un changement perceptible dans la forme de l’ombelle qui évolue vers une forme convexe, via des changements au niveau des rayons de l’ombelle.

Ombelles à peine fructifiées et déjà en cours de repliement 
La fanaison des fleurs fécondées fait ressortir l’ovaire qui se trouvait sous la fleur (position dite infère) : sa structure double originelle transparaît dans le fruit vert formé de deux éléments jumelés, les méricarpes (voir la chronique générale sur les ombellifères) ; ils conservent à leur sommet chacun un style, comme une petite corne. Chaque méricarpe ne renferme qu’une seule graine et correspond donc à un fruit sec à une seule graine soudée à la paroi du fruit, un akène. On peut donc assimiler, d’un point de vue fonctionnel, chacun des méricarpes à un fruit- graine. D’ailleurs quand vous achetez un sachet de graines de carottes cultivées, en fait ce sont des akènes donc des fruits que vous semez.

Nids d’oiseaux

Avec la maturation de ces fruits-graines, on assiste à un fort repli des ombelles sur elles-mêmes, mouvement accompagné par l’involucre (voir la chronique) sous la base des rayons de l’ombelle : les rayons s’épaississent et se courbent fortement vers l’intérieur, entraînant avec eux les ombellules de fruits vers le centre. Les ombelles fructifiées, garnies de fruits-graines verts pour l’instant, prennent ainsi une forme de boule compacte ou d’entonnoir un peu ouvert au sommet, surnommée nid d’oiseau ou nid d’abeille en langage populaire. Ce comportement diffère radicalement de ce qui se passe chez la majorité des ombellifères où les ombelles fructifiées restent largement étalées.
Dans la foulée, les fruits-graines entament leur maturation en séchant et en virant au brun ; la tige porteuse et l’ombelle entière sèchent à leur tour mais conservent leur port replié en nid d’oiseau. Quelques semaines plus tard, la fructification des ombelles latérales terminée à son tour, toute la plante sèche sur pied, comportement typique d’une plante bisannuelle (voir la chronique sur le cycle) ; nous sommes alors juste en fin d’été/début d’automne. Les peuplements denses de carottes sauvages prennent un aspect radicalement différent, loin des tapis de petits nuages blancs des ombelles fleuries : on se trouve devant un fouillis de boules compactes entourées de leurs rayons repliés.



Avec la mort (programmée) de la plante, commencent de surprenants mouvements d’ouverture ou au contraire de fermeture encore plus étroite de l’ombelle fructifiée. Les rayons externes répondent aux changements d’humidité dans l’air (hygroscopie) en se tordant vers l’axe central de l’ombelle quand l’humidité augmente. Ainsi par temps sec, les fruits-graines se trouvent exposés au vent et facilement arrachés et dispersés. Au contraire par temps humide, les ombelles se ferment et ainsi, les fruits-graines secs « enfermés » ne peuvent plu être dispersés (voir ci-dessous). Ce processus peut s’étaler sur tout l’hiver puisque les plantes sèches sur pied persistent jusqu’à la fin de l’hiver (voir la chronique sur le cycle) : ainsi, la dispersion se fait d manière discontinue et étalée dans le temps ce qui augmente les chances de rencontrer des conditions nouvelles ; la carotte « ne jette pas ses oeufs de son panier d’un seul coup » en somme.


Ces mouvements hygroscopiques s’expliquent par une orientation différentielle de microfibrilles dans les cellules des faces externes/internes des rayons primaires : elles sont orientées majoritairement de manière transversale dans la moitié externe du rayon versus en long dans la moitié interne. Le jeu contradictoire de raccourcissement des fibrilles sous l’effet du dessèchement se traduit par des déformations réversibles des rayons.

Fluctuante

A maturité, les deux méricarpes d’abord collés s’écartent et se séparent, chacun porté au bout de l’une des deux extrémités du porte-fruits fourchu ; ils vont donc désormais suivre chacun leur voie de manière indépendante. La saison de libération des fruits-graines commence à partir de fin août et dure jusqu’au printemps suivant, tant que les plantes sèches restent sur pied. Le décrochement des fruits du court filament qui les porte commence par ceux les plus à l’extérieur puis se propage progressivement vers l’intérieur. Ainsi, au cœur de l’hiver, on trouve souvent des ombelles dégarnies à la périphérie mais encore chargées de fruits au centre.

Octobre 
La dispersion de l’ensemble des fruits-graines d’une ombelle donnée peut s’étaler sur plusieurs mois selon les épisodes d’ouverture des ombelles nids d’oiseaux (voir ci-dessus). Mais il existe de fortes disparités : certaines plantes se débarrassent de leurs fruits en moins d’un mois alors que d’autres les retiennent presque tout l’hiver. Divers facteurs influent sur la libération des graines : par exemple, la qualité de l’automne (en termes de précipitations) qui suit la production sélectionne une dispersion soit graduelle, soit rapide.

Les populations pionnières récemment installées (voir la chronique sur les milieux de vie) tendent à libérer plus tôt leurs graines : leur croissance est plus rapide dans les milieux fraîchement ouverts. Inversement, dans les peuplements plus anciens où la végétation environnante commence à concurrencer les carottes, une forte proportion de graines sera libérée plus tard au cours de l’hiver. Or, le sort des graines varie selon le moment de leur libération. En collectant des graines de plantes ayant fleuri à des moments différents, on constate que la croissance est plus forte pour les plantules issues de graines libérées tôt en saison. Elles proviennent de plantes ayant fleuri plus tôt et donc bénéficié a priori de conditions plus favorables autorisant la production de fruits de meilleure qualité. Ainsi, les jeunes peuplements dans des milieux ouverts très favorables verront leurs populations se renforcer ; inversement, dans les peuplements anciens, plus de graines libérées en hiver auront de chances d’être dispersées au loin (voir ci-dessous) et de coloniser ainsi de nouveaux milieux favorables.

Les fruits des ombellules externes tombent en premier ; le reste est ici renfermé au coeur du « nid » 
Nid ouvert en hiver
Ces conditions très changeantes entretiennent une forte variabilité dans la dispersion des graines ce qui augmente les chances de rencontrer un contexte favorable. Elle dépend directement des mouvements hygroscopiques des nids d’oiseaux dont on pense qu’ils seraient, pour une part, sous contrôle génétique. Le fait que les ombelles latérales d’une plante donnée se comportent comme l’ombelle terminale tend à conforter cette hypothèse. Cette remarquable plasticité permet à la carotte sauvage de s’adapter à des milieux de vie qui évoluent très vite ou apparaissent brutalement.
Nids de … prédateurs ?

On pourrait aussi penser que la formation des nids d’oiseaux procure aux fruits de la carotte une certaine protection contre les prédateurs de graines en les « renfermant » de manière relative. En pratique, on constate que ces nids fermés servent très souvent d’abri à divers insectes dont une forte majorité de punaises pentatomidés de diverses espèces ; elles se tiennent souvent au bord des ombelles contractées et se réfugient à l’intérieur à la moindre alerte ; lors d’épisodes de fortes chaleurs, elles se réfugient aussi dedans pour se mettre à l’abri des rayons du soleil brûlant.

Graphosome rayé : l’espèce la plus commune sur les carottes 

Les graphosomes rayés (voir la chronique) sont de loin les plus courants, parfois en groupes sur une même infrutescence ; on observe aussi des punaises des baies, des punaises potagères, des punaises à pointes, … Or, ces insectes végétariens se nourrissent essentiellement de sucs de graines encore vertes qu’ils percent avec leur rostre piqueur-suceur pour en aspirer le contenu (voir la chronique) ; on peut les voir d’ailleurs attablés à l’extérieur des nids d’oiseaux en train de siroter le contenu des graines. Autrement dit, ces ombelles refermées ressemblent plus à des « pièges » pour la carotte sauvage en favorisant la présence de ses ennemis ?

Punaise à cornes 
Punaise des baies
Aux USA où la carotte sauvage a été introduite et est devenue invasive, on a étudié sa relation avec une punaise Lygéidé locale (Lygus superbus) qui l’a adopté comme plante-hôte. On a montré que la prédation de ces punaises réduit la production du nombre de graines et diminue la capacité de germination des graines ponctionnées et affecte ensuite la croissance des jeunes plantules. Par contre, dans ce cas, cette punaise se comporte aussi en prédateur d’insectes ce qui peut procurer quelques bénéfices pour la carotte ; mais ce n’est pas le cas des punaises pentatomidés, strictement végétariennes. En se promenant sur les inflorescences, ces punaises peuvent incidemment participer un peu à la pollinisation (voir la chronique) et apporter donc un plus relatif. Au final, il n’est donc pas sûr qu’elles affectent considérablement le potentiel reproducteur considérable de la carotte.

Eclosion d’oeufs de punaises 
Lieu de rendez-vous et d’accouplements 
Abri pour jeunes punaises
On observe aussi de jeunes punaises qui s’abritent dans les nids d’oiseaux et même des pontes fixées sur les ombelles. En tout cas, par ce biais, les carottes sauvages participent au maintien d’un autre pan de biodiversité animale, en sus de leur rôle de corne d’abondance pour pollinisateurs (voir la chronique).
On peut observer plus rarement d’autres visiteurs dans ces nids qui se comportent avant tout en commensaux cherchant un abri sans être pour autant des consommateurs ; ainsi, il m’est arrivé d’observer des groupes de Ténébrionidés (Coléoptères) réfugiés dans ces nids ou bien un grillon d’Italie, bien calé la tête en bas dans la position de repos typique de nombreux orthoptères.

Grillon d’Italie caché dans un nid d’oiseau que j’ai entrouvert pour faire la photo.
Accrocheuse

Les fruits des carottes ne dépassent pas 2 à 4mm de long. De forme ovale, ils portent 5 côtes primaires avec des cils ou poils courts, intercalées avec 4 côtes secondaires plus grandes, ailées et dotées de forts aiguillons. Sur un fruit double encore non séparé, on a donc ainsi 8 rangées d’aiguillons raides, fins, libres entre eux à leur base.


Rangée de poils raides alternant avec les rangs d’épines
Ils se terminent chacun à leur sommet par des barbes recourbées au nombre variable allant de 1 à 5 ; selon leur nombre, leur angle d’orientation par rapport à l’épine varie : moins de 90° quand ces barbes sont 4 à 5, environ 90° pour 2 à 3 et plus de 90° pour une barbe seule.
Barbes au bout des épines 
Encore plus nettes sur des fruits secs
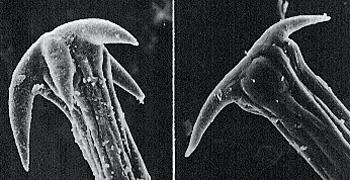
Une telle structure suggère immédiatement un mode dispersion bien connu par accrochage au pelage d’animaux passant à proximité ou sur les vêtements d’humains : c’est le processus d’épizoochorie (voir la chronique).
Des expériences sur des peaux naturalisées montrent que les fourrures fines (par exemple lapins et souris) n’accrochent pas ou peu ces fruits-graines ; par contre les fourrures touffues (par exemple raton-laveur) les retiennent bien et les fruits ne s’enlèvent que si on les extrait. Si on frotte une tête fructifiée sèche sur un vêtement, un certain nombre de fruits s’accrochent mais sans être ancrés très fortement sauf sur de la laine.

En Espagne, on a réalisé une expérience grandeur nature sur des moutons qui effectuent chaque année une transhumance d’un mois en parcourant 4000 km (voir la chronique sur cet exemple) ; on a placé des graines de carotte (entre autres) sur certains moutons ; à l’arrivée, on tond ces moutons et on compte le nombre de graines restantes : 7% dans le cas de la carotte. Même si ce chiffre reste modeste, il ouvre néanmoins d’extraordinaires possibilités de voyage à très long cours pour les fruits sachant qu’un certain nombre ont du se décrocher progressivement au fil du voyage.
Des mammifères herbivores peuvent aussi consommer des graines en broutant dans les prairies : on a ainsi réussi à faire germer des graines retrouvées dans des excréments de chevreuil ou du crottin de cheval. Là, on se trouve dans un scénario d’endozoochorie, i.e. de voyage « à l’intérieur » (voir la chronique) ; si les animaux se déplacent après la consommation le temps du transit intestinal, ils vont déposer ces graines à une certaine distance.
De manière anecdotique, on a aussi retrouvé des graines incorporées dans la structure de nids d’oiseaux soit une autre forme indirecte de transport.
Diplochore ?
Néanmoins, dans de nombreux milieux habités par la carotte sauvage, on se dit qu’il ne doit pas passer tant d’animaux susceptibles de prendre en charge des fruits et qu’il doit donc exister au moins un autre mode de dispersion moins évident. De plus, quand on se promène dans un champ de carottes fructifiées, on ne récupère que peu de fruits sur ses vêtements alors que la production est colossale (jusqu’à 15000 fruits/pied). C’est le raisonnement tenu par les scientifiques aux USA où la carotte sauvage prospère notamment dans les grandes plaines cultivées : les épines et ailes des fruits ne freineraient-ils pas leur chute dans l’air et ne favoriseraient-ils pas ainsi le transport par le vent (anémochorie : voir la chronique) ? Une série d’expériences a confirmé cette hypothèse en situation ouverte où le vent peut s’exprimer : les fruits aux épines « tondues » parcourent une distance moyenne inférieure à celle des fruits intacts (0,33m versus 0,45m) ; les fruits dépourvus de barbes au bout des épines (voir ci-dessus) vont eux aussi un peu moins loin : 0,49m versus 0,53m. donc, épines et barbes apportent bien un léger plus favorisant la dispersion par le vent mais à courte distance en moyenne.
Cependant, toujours aux USA, dans les grands espaces ouverts, un autre élément intervient en hiver : la couche neigeuse ; une fois tombés sur la neige gelée, les fruits peuvent être de nouveau mobilisés et déplacés par le vent : des expériences donnent des distances moyennes parcourues ainsi de 2 à 2,50m avec un maximum de 6,25m pour un vent à 5-6 km/h.
Autrement dit, dans certains environnements (avec du froid et du vent en hiver), ce mode de dispersion peut largement concourir au transport à distance des fruits et vient compléter l’épizoochorie. On parle de diplochorie, i.e. de deux modes de dispersion qui ne s’excluent pas mutuellement. Peut-être qu’aux USA le second mode prédomine compte tenu du contexte climatique et des milieux alors que dans nos pays, ce serait plutôt le premier.
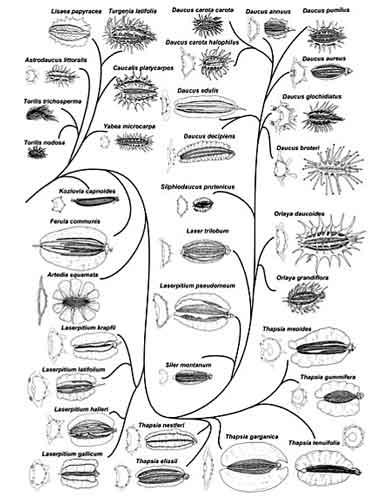
La comparaison de la carotte avec ses plus proches parents au sein de leur famille apporte un éclairage complémentaire : elle se place dans la tribu des Scandicae où le trait ancestral prédominant est la présence de côtes marquées sur les fruits développées en ailes facilitant l’anémochorie. Seules les lignées plus récentes des carottes (Daucinae) et des torilis (Torilidinae) possèdent des fruits épineux, soit un caractère dérivé apparu ultérieurement. D’ailleurs, on a été amené à reclasser dans le genre carotte (Daucus) une espèce (D. annuus) ayant des fruits dotés d’appendices intermédiaires entre ailes et épines : elle vit sur les îles de Macaronésie dépourvues de mammifères indigènes et aurait donc évolué récemment par réversion vers le trait ancestral « ailes sans épines ».

Torilis des champs à fruits épineux 

Laser à feuilles larges à fruits ailés 
Retour aux nids
Cette dispersion par le vent dépend par ailleurs des mouvements des rayons de l’ombelle à l’origine des nids d’oiseaux évoqués ci-dessus. Ces mouvements (relèvement ou rabaissement) ont lieu en fonction d’une large gamme d’humidité et s’amplifient avec l’âge, i.e. au cours de l’hiver sur la plante sèche. Pour tester le rôle de ces mouvements, on souffle sur des ombelles fructifiées plus ou moins contractées ou étalées et on évalue la distance moyenne de dispersion. Résultat : le vent transporte sur une plus grande distance en moyenne les fruits arrachés à des ombelles très relevées par rapport à celles très étalées ; le transport se fait sur de courtes distances pour une humidité faible et sur de plus longues distances pour une humidité élevée. Ce couplage dispersion par le vent/mouvements hygroscopiques assure ainsi que la descendance sera dispersée à des distances très variées depuis la plante mère et qu’au final, tous les fruits finiront par être dispersés compte tenu de la durée de « vie » des plantes sèches et de la poursuite des mouvements.
Tout ceci créé une mosaïque de conditions étalée sur tout l’hiver qui augmente considérablement la probabilité que les fruits soient transportés plus ou moins loin et atteignent peut-être de nouveaux milieux sous des conditions météorologiques favorables à la germination. La stratégie de la carotte repose avant tout sur cette capacité de dispersion et d’atteindre vite de nouveaux milieux ouverts favorables car très peu de graines persistent au-delà de 1 à 2 ans : elle ne dispose donc que d’une banque de graines à court terme ; et encore, la prédation par les rongeurs reste limitée du fait de la petite taille des fruits. Ainsi, elle ne peut compter sur le « réveil » des graines des dizaines d’années après leur arrivée au sol à la faveur d’une perturbation comme par exemple les coquelicots (voir la chronique).
Bibliographie
Seed dispersal in wild carrot (Daucus carota). E.P. Lacey The Michigan botanist p. 15-20. 1981
THE ANATOMICAL BASIS FOR HYGROSCOPIC MOVEMENT IN PRIMARY RAYS OF DAUCUS CAROTA SSP. CAROTA (APIACEAE) ELIZABETH P. LACEY, et al. BOT. GAz. 144(3):371-375. 1983.
The Influence of Hygroscopic Movement on SeedDispersal in Daucus carota (Apiaceae) E. P. Lacey Oecologia 47, 110-114(1980)
Extreme long-distance seed dispersal via sheep. Pablo Manzano and Juan E Malo. Front Ecol Environ 2006; 4(5): 244–248
Evolutionary shifts in fruit dispersal syndromes in Apiaceae tribe Scandiceae Aneta Wojewódzka et al. Plant Systematics and Evolution (2019) 305:401–414
Timing of Seed Dispersal in Daucus Carota Elizabeth P. Lacey Oikos, Vol. 39, No. 1. (1982), pp. 83-91.
