Les rapaces ne sont plus ce qu’ils étaient
Le terme de rapace, très usité dans le langage courant, désigne (d’après la définition du Robert) « un oiseau carnivore aux doigts armés d’ongles forts et crochus (serres), au bec puissant, arqué ou pointu » ; ce sont les oiseaux de proie. La culture populaire en fait un groupe en soi, le partageant en deux sous-groupes, les rapaces diurnes et les rapaces nocturnes. Si cette classification informelle a longtemps été aussi celle adoptée par le monde scientifique, elle vient au cours des dernières décennies de littéralement exploser sous les coups de boutoir des phylogénies moléculaires successives (2,3 et 4), i.e. la comparaison des séquences génétiques des différentes espèces permettant de reconstituer leur histoire évolutive. Désormais, on ne peut plus parler que de rapaces nocturnes comme groupe évolutif partageant un ancêtre commun alors que les rapaces diurnes se trouvent répartis dans plusieurs groupes non directement apparentés de la classification des oiseaux.
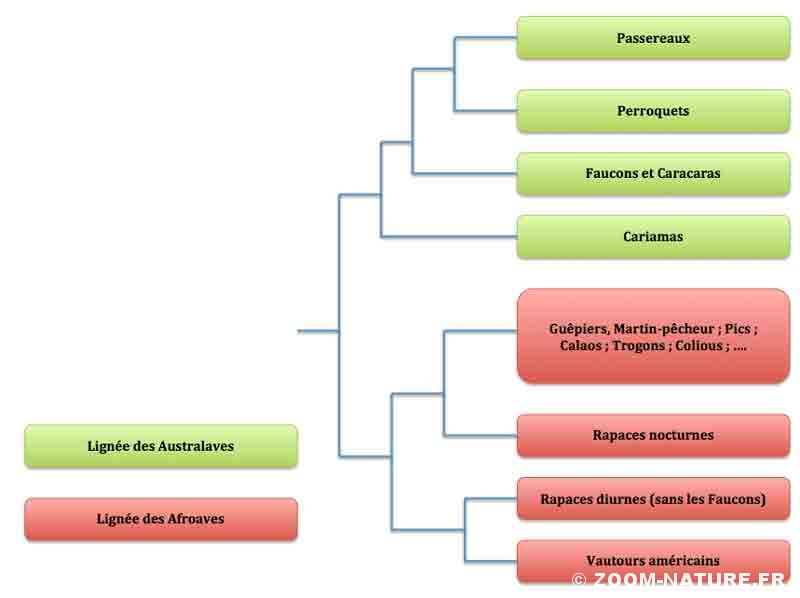
Arbre phylogénétique du groupe d’oiseaux terrestres dans lequel se retrouvent les différents groupes de Rapaces
Les rapaces nocturnes
Les Strigiformes, le nom scientifique pour les Rapaces nocturnes, forment effectivement un groupe évolutif clairement défini par un ensemble de caractères partagés uniques (des synapomorphies : voir la chronique sur les signatures osseuses des dinosaures) dont : une « arcade sourcilière » prononcée dû au développement d’une excroissance de l’os frontal ; des trous auditifs (les oiseaux n’ont pas d’oreille externe ou pavillon) qui ne sont pas placés à la même hauteur (donc asymétriques) qui leur procure une audition en « stéréo » très performante ; une disposition particulière des doigts : le 4ème doigt, ordinairement orienté vers l’avant, est suffisamment mobile pour se tourner vers l’arrière en compagnie du doigt n°1 classiquement tourné en arrière ce qui donne un port en croix typique ; des parties du crâne sont épaissies par de l’os spongieux. On distingue deux familles au sein de cet ordre : les effraies et les phodiles (Tytonidés) avec leur disque facial en forme de cœur sur une face aplatie et les autres « chouettes et hiboux » (Strigidés).
-

- Pied de rapace nocturne avec les serres

-

- Hibou Grand-duc (Zoo du Pal 03). La distinction chouette/hibou ne relève pas de la classification mais d’un usage populaire de ces mots

-

- Disque facial en forme de coeur d’une effraie des clochers (Muséum H. Lecoq ; Clermont-Ferrand)

Si la délimitation du groupe ne pose pas de problème, sa position dans l’arbre des oiseaux reste toujours sujette à discussion. On l’a placé comme groupe-frère (le plus proche parent) des accipitriformes (une partie des rapaces diurnes : voir la suite), l’ensemble des deux étant alors connu sous l’appellation de Raptores, ce qui restaurait en quelque sorte l’idée de « rapaces » réunis. Des données plus récentes tendent à ne plus les placer aux côtés des Accipitriformes mais sur une autre branche, comme groupe frère (peut-être ?) des Colious, les curieux « oiseaux-souris » qui ressemblent très superficiellement à des perruches.
Les faucons ne font partie des Rapaces !
Eh oui, il va falloir s’y faire car toutes les phylogénies récentes concordent sur ce point : le groupe des faucons et des caracaras (famille des Falconidés) n’est pas du tout apparenté aux autres « rapaces diurnes » mais se place sur une branche nettement divergente, celle des Australaves aux côtés des Cariamas d’une part (voir la chronique consacrée à ces oiseaux très particuliers) et des Psitacciformes (perroquets, perruches et cacatoès) et des Passériformes, l’immense groupe des Passereaux. Autrement dit, les faucons et caracaras sont plus proches parents des perroquets ou des passereaux que des buses ou des aigles ! Pas grand chose a priori dans leurs apparences ne permettait se suspecter une telle absence de parenté directe avec les autres Rapaces diurnes à part trois caractéristiques directement observables : un bec crochu avec une dent (dite tomiale) latérale ; des griffes (serres) pratiquement égales et relativement moins courbées, sur des doigts assez fins en lien avec une méthode de capture et d’immobilisation des proies différente (voir la chronique sur les serres des rapaces qui développe longuement cet aspect très intéressant) ; des œufs avec une coquille peu épaisse ce qui leur a valu des déboires suite à l’intoxication par certains pesticides comme le D.D.T. qui réduisaient encore plus l’épaisseur et provoquaient des pertes considérables lors de l’incubation.
-

- Faucon pèlerin (Muséum de Bourges)

-

- Les caracaras sont aussi classés avec les faucons dans l’ordre des Falconiformes (zoo du Pal; 03)

-

- Cariama huppé : l’un des plus proches parents actuels des Faucons

-

- Les Psittaciformes (ici des Aras au zoo des sables d’Olonne) sont l’autre groupe actuel lenplus proche parent des Falconiformes.

Exit donc l’ancien groupe des Falconiformes dans lequel on plaçait tous les rapaces diurnes, en prenant au passage comme « bannière » les faucons, les moins « rapaces » d’entre tous ! Bienvenue aux « nouveaux » Falconiformes, réduits aux seuls faucons et caracaras. On notera néanmoins qu’ils se situent « quand même » dans la même grande lignée d’oiseaux terrestres que les autres rapaces mais dans une lignée qui a divergé très tôt.
R.J. Monneret, grand spécialiste des faucons et de leur protection en France, fait remarquer qu’il n’est pas surpris de la parenté perroquets/faucons autant dans leur comportement que leur façon de voler. Par contre, il s’étonne de la « proximité » avec le passereaux du fait d’un trait typique : « une des différences remarquables entre faucons et passereaux est le fait que pour se gratter la tête les passereaux passent leurs pattes par dessus l’aile, alors que les faucons, comme les autres « rapaces », se grattent directement en passant devant« . Cette fine observation ne me semble pas pour autant incompatible vu que les faucons en tant que groupe occupent une position plus ancienne ; ce comportement a du apparaître secondairement dans la lignée des passereaux sous réserve qu’il n’existe pas dans d’autres lignées antérieures (à vérifier !).
Le cas des vautours américains
Il y a encore quelques décennies, l’un des grands sujets de discussion concernait la place des « vautours du Nouveau monde », les Cathartidés regroupant condors, urubus et vautour pape. Une étude basée sur une méthode aujourd’hui abandonnée (hybridation des ADN) car trop imprécise en avait fait le groupe frère des …. Cigognes ! Ils partageaient notamment une curieuse habitude, celle de déféquer sur leurs tarses pour faciliter la déperdition de chaleur ou d’avoir des narines perforées (de profil, on voit « à travers le bec »). Depuis, les analyses moléculaires ont infirmé complètement cette hypothèse qui a persisté un temps pour les placer au plus près des autres rapaces diurnes (donc sans les faucons) soit dans un ordre à part (Cathartiformes ; option retenue sur l’arbre proposé ci-dessus), soit comme simple famille au sein de l’ordre des Accipitriformes selon l’importance relative que l’on accorde à leurs originalités. En effet, ils possèdent quand même des caractères un peu particuliers comme le bec assez court, les narines perforées, la tête petite entièrement dénudée et vivement colorée, les doigts grêles aux griffes courtes. Ils partagent avec les « vautours de l’Ancien Monde » le régime de charognards mais les deux sexes sont de même taille contrairement à ce qui se passe chez ces derniers (sauf chez le grand condor où les mâles sont nettement plus gros que les femelles : remarque de R.J. Monneret).
-

- Le vautour pape d’Amérique du sud est un membre de la famille des Cathartidés, souvent élevée au rang d’ordre (Cathartiformes)

Les « vrais » rapaces diurnes
Il nous reste donc, réunis ensemble, tous les autres Rapaces diurnes dans l’ordre des Accipitriformes.
-
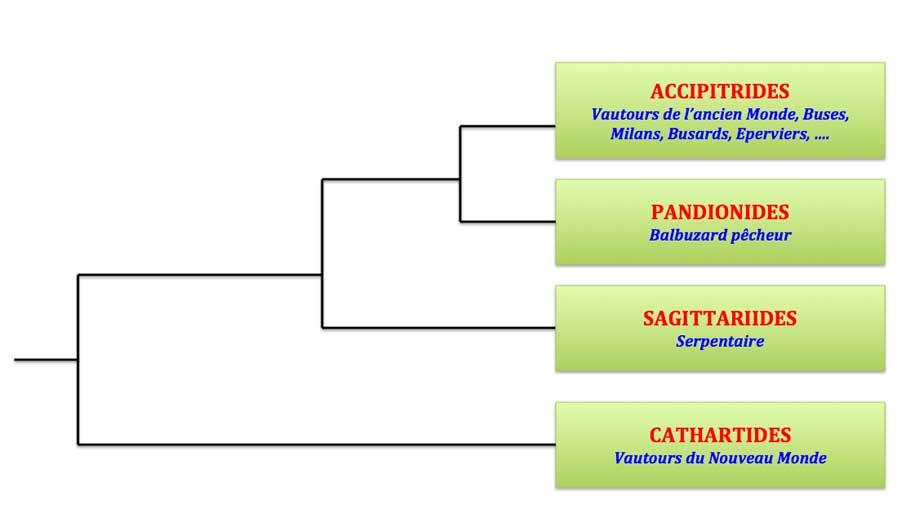
- Arbre phylogénétique de l’ordre des Accipitriformes. Ici, les Cathartidés ont été interprétés comme une famille et non un ordre (ce qui ne change rien aux parentés entre groupes).
On distingue trois familles :
– les Accipitridés : vautours de l’Ancien Monde (dont les gypaètes), buses, milans, busards, circaètes, pygargues, élanions, « aigles », éperviers et autours, bondrées, harpies, … et bien d’autres puisque cette famille compte 248 espèces réparties dans 69 genres. Une belle diversité donc !
– les Sagittariidés qui ne comprennent qu’une seule espèce, le très étrange secrétaire ou sagittaire des savanes africaines, oiseau atypique à plus d’un titre ; cette famille se situe à la base de l’arbre
– les Pandionidés avec « une » espèce bien connue, le balbuzard pêcheur, rapace spécialisé dans le régime piscivore (voir la chronique consacrée au balbuzard à propos du nombre réel d’espèces) et qui possède des caractères uniques par rapport aux autres rapaces de son groupe (voir la chronique sur la famille des Pandionidés).
-

- Vautour fauve en vol, un exemple de Vautours de l’ancien Monde, non directement apparentés aux Cathartidés, les vautours du Nouveau Monde

-

- Le percnoptère d’Egypte (présent en France) est un autre vautour dans la famille des Accipitridés. Zoo du Pal ; 03

-

- Milan royal en vol (Accipitridés). Photo J. Lombardy

-

- Bondrée apivore (Accipitridé). Photo J. Lombardy

-

- Le Balbuzard pêcheur, rapace piscivore très spécialisé est classé dans une famille à part, les Pandionidés. Photo J. Lombardy

-

- Aigle pêcheur d’Afrique (Accipitridé), lui aussi en partie piscivore mais moins spécialisé que le Balbuzard (Photo D . Bermudez)

Des convergences à la pelle !
Si on s’attarde sur les caractères propres au groupe des Accipitriformes (y compris les cathartidés), on trouve plusieurs caractères osseux originaux comme l’os du pubis qui se trouve fusionné au bout du bassin dont l’extrémité s’incurve fortement (ce qui donne aux rapaces ce port « vertical » au repos) et une arcade sourcilière très développée (qui donne cet air guerrier aux rapaces ou méchant comme disent les chasseurs !) issue d’une excroissance de l’os préfrontal. Mais si l’on consulte les caractères propres aux Falconiformes (dans leur définition actuelle), on découvre …. les mêmes caractères ! Supercherie diront certains esprits mal intentionnés ! Il s’agit simplement d’un bel exemple de convergence évolutive où, dans deux lignées relativement éloignées mais partageant un mode de vie prédateur commun, les mêmes adaptations anatomiques apparaissent indépendamment. Ceci s’explique par le jeu des gènes du développement communs aux oiseaux de cette grande lignée (Australaves et Afroaves réunis) qui ont du subir dans ces deux branches des mutations ayant subi la même pression de sélection.
On notera aussi que des convergences existent aussi avec les Strigiformes, les Rapaces nocturnes avec notamment le développement de serres (mais assez différentes) et d’une arcade sourcilière mais formée à partir d’un os différent ; autrement dit, là, on peut débusquer la convergence assez facilement ! Enfin, on a aussi une nette convergence entre les Vautours de l’Ancien monde (classés dans les Accipitridés) et ceux du Nouveau Monde (les Cathartiformes), là encore sous la pression sélective d’un mode de vie commun, le régime charognard ou nécrophage.
Nous verrons dans la chronique sur les serres qu’il y a bien de réelles différences « profondes » entre faucons et autres rapaces justement dans le mode de chasse et d’immobilisation des proies. Différences subtiles mais différences éclairantes néanmoins qui permettent même de faire des rapprochements entre les faucons et leurs parents perroquets ou passereaux ! Ceci montre que la phylogénie moléculaire (et souvent décriée à tort par les naturalistes comme abstraite) nous apporte des éclairages qui orientent nos observations et nous amènent à reconsidérer nos points de vue et à voir « autrement » des oiseaux pourtant familiers (pour les ornithologues au moins !).
BIBLIOGRAPHIE
- Bird families of the world. D.W. Winkler et al. Lynx Ed. 2015
- A Phylogeny of Birds Based on Over 1,500 Loci Collected by Target Enrichment and High-Throughput Sequencing. John E. McCormack, Michael G. Harvey, Brant C. Faircloth, Nicholas G. Crawford, Travis C. Glenn, Robb T. Brumfield. PLOS ONE. January 2013 | Volume 8 | Issue 1
- A Phylogenomic Study of Birds Reveals Their Evolutionary History Shannon J. Hackett, et al. Science 320, 1763 (2008)
- Whole-genome analyses resolve early branches in the tree of life of modern birds. Jarvis et al. Science. 2014 December 12; 346(6215): 1320–1331.
A retrouver dans nos ouvrages
Page(s) : p. 461-463

Page(s) : p. 479-480
Page(s) : p. 464-465
