Magnolias for ever
Magnolia grandiflora

Parmi les arbres ornementaux cultivés sous nos climats, le magnolia à grandes fleurs se détache par l’opulence et la beauté remarquables de ses énormes fleurs blanc pur. Personnellement, j’adore ces fleurs d’autant que j’ai eu la chance de m’en voir offrir deux jeunes pieds que j’ai plantés juste devant mes fenêtres ; ainsi, je peux suivre en été, heure par heure, tout le processus de la floraison qui se répète des dizaines de fois sur le même arbre. Vu la taille des fleurs, nul besoin de loupe pour observer et suivre le processus qui s’étale sur trois à quatre jours par fleur ; par contre, on réalise vite qu’il se passe plein de choses « en coulisses » compte tenu des transformations profondes qui affectent cette fleur sur cette courte période. Quels secrets se cachent derrière ce déploiement fastueux lors de la floraison ?
Fleur primitive ?
-

- Le magnolia à grandes fleurs est devenu un arbre ornemental très répandu et apprécié (Jardin des Plantes : Paris)

La réputation de fleurs primitives, presque des fossiles vivants, colle aux Magnolias depuis longtemps ; cela tient d’une part à leur grande taille, à la disposition des pièces florales et surtout au fait que la famille des Magnoliacées fait partie d’une lignée très ancienne, une des plus anciennes qui s’est détachée à la base de l’arbre des plantes à fleurs. De ce fait, on prétendait que leurs fleurs correspondaient au type ancestral des premières plantes à fleur. Or, si l’on examine le riche registre fossile de cette famille et de ses proches, on y découvre en fait de nombreuses formes avec de petites fleurs !
On disait aussi que les fleurs des Magnolias étaient pollinisées essentiellement par des scarabées qui en déambulant au milieu de ces grosses fleurs en coupe transportaient de manière désordonnée du pollen ; on prétendait que les abeilles et bourdons (hyménoptères) visitaient peu ces fleurs où elles se sentaient mal faute de guides visuels. Or, une série d’études récentes apportent une toute autre image de ces arbres et arbustes : la majorité possèdent en fait des fleurs très spécialisées dotées de dispositifs d’attraction (dont des parfums) et de guidage, de capacités de mouvements internes au sein des fleurs, de sources de nourriture variées et spécialisées, … Bref, on est loin de la fleur primitive qui serait restée figée « dans son jus ancien » !
Cependant, la grande difficulté pour aborder ce sujet reste la grande diversité du genre Magnolia (en l’élargissant aux « petits » genres satellites très proches) avec plus de 230 espèces réparties de manière relativement disjointe dans le monde : un groupe en Asie orientale avec des isolats au Japon et un groupe centré sur l’Amérique du nord jusqu’au Mexique. Donc, des histoires différentes et des contextes différents si bien que d’une espèce à l’autre on trouve des variations considérables pour un élément donné de la fleur. Ici, nous allons donc nous concentrer sur l’exemple du magnolia à grandes fleurs en sachant donc qu’il ne représente qu’une partie de la diversité du genre.
Spirales et verticilles
La structure et le développement de cette fleur dans le boutons apportent quelques surprises par rapport à la vision classique que nous avons des fleurs en général. Le bouton floral se trouve enveloppé dans une bractée (une spathe) et non pas dans un calice (une feuille modifiée), caractère inhabituel qui a renforcé l’image de fleur primitive ancestrale ! De calice en fait, il n’y en pas vraiment puisque les pièces florales colorées (blanc pur dans le cas de ce magnolia) normalement au nombre de neuf sont toutes identiques : on parle donc de tépales et non de pétales (corolle) et de sépales (calice). Cependant, la mise en place de ces pièces se fait bien en deux temps : d’abord un premier cercle de trois (soit l’équivalent d’un calice mais non différencié) puis successivement deux autres cercles de trois vers l’intérieur (soit la corolle). En fait de cercle (verticille en botanique), si on regarde de très près les deux derniers, on se rend compte qu’ils ne se trouvent pas exactement au même niveau et ne sont pas espacés régulièrement ; en fait, ces six tépales internes seraient initiés selon une spirale très serrée.
-

- Fleur en bouton enveloppée dans une spathe qui correspond à une bractée

-

- A l’éclosion, la bractée tombe très vite et laisse apparaître les tépales blanc pur

Les pièces fertiles (étamines et pistils) de la fleur vont s’organiser autour d’un axe allongé central qui trône au centre de la fleur. Sur une tige rougeâtre cylindrique à la base, presque en même temps que le dernier cercle de tépales, les étamines se forment selon des lignes spirales bien visibles une fois qu’elles sont tombées. Enfin, quand toutes les étamines sont initiées, sur la massue terminale de l’axe, apparaissent, là encore selon des lignes spirales, des rangées de pistils globuleux terminés par un stigmate crochu.
-
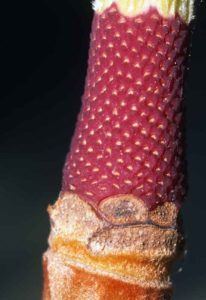
- Base rouge de l’axe central qui portait les étamines : on devine leur insertion le long de lignes spirales
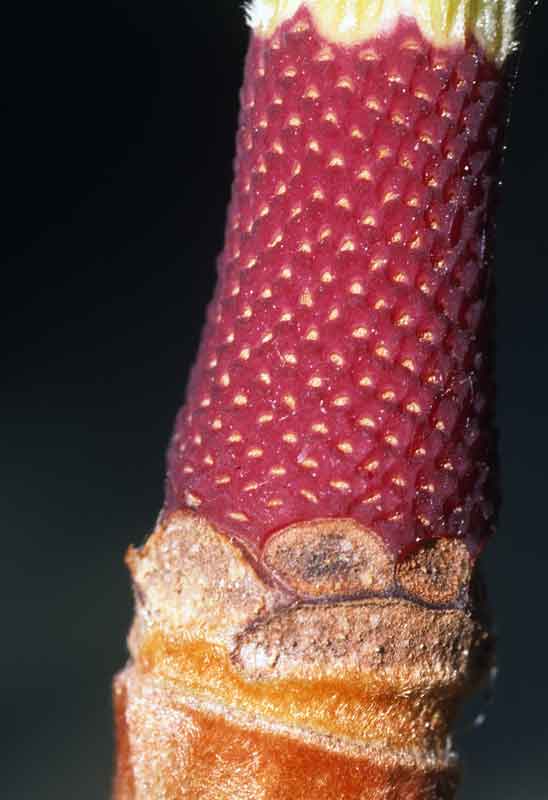
Ainsi globalement, cette fleur s’organise avant tout selon un schéma spiralé qui va transparaître le plus au niveau des pistils à maturité, quand ils seront devenus des fruits. On suppose que la disposition verticillée qui se dessine pour les tépales est dérivée de la disposition ancestrale spiralée qui perdure au niveau des pièces fertiles ; dans les lignées plus récentes, la disposition verticillée deviendra prépondérante.
Tépales
Revenons donc au « premier cercle » celui des neuf tépales ; l’ensemble disposé en coupe évasée rappelle une énorme tulipe dont le diamètre peut atteindre 30cm ! Ceci vai-ut aux Magnolias d’être régulièrement surnommés des tulipiers mais ce nom désigne aussi deux autres arbres du genre Liriodendron (dont le tulipier de Virginie) qui sont les plus proches parents des Magnolias au sein de la famille des Magnoliacées. Chaque tépale d’un blanc nacré superbe possède une consistance charnue et cireuse ; à la fanaison, ils se rétractent et prennent une coloration rouille qui, de mon point de vue, ne manque pas d’une certaine esthétique en tant que mémoire d’une beauté fugitive. En effet, chaque fleur ne dure que trois à quatre jours à partir de son éclosion avant de faner rapidement. Chaque arbre produit de nombreuses fleurs mais très étalées sur plusieurs semaines ou mois à partir de la mi- juin (sous nos climats). Ainsi, chaque jour de l’été, sur un vieil arbre, on peut trouver des fleurs à différents stades !
-

- Tépales en fin de floraison prenant un aspect de vieux cuir !

Chez cet arbre à croissance lente, les premières floraisons ne se produisent qu’à un âge un peu avancé : chez des arbres issus de semis de graines, il faudra attendre en moyenne 25 ans alors que chez des arbres issus de greffes ou de marcottage de branches basses, ce délai s’abaisse à 15 ans en moyenne. D’ailleurs, historiquement, le premier magnolia à grandes fleurs fut rapporté en France (depuis le sud-est des USA, son pays natal) en 1711 et planté près de Nantes ; il connut un piètre succès jusqu’en 1732 où il se mit à fleurir et devint alors un objet d’émerveillement et d’engouement horticole !

Androcée
L’androcée (l’ensemble des étamines) occupe la base de l’axe central et forme un gros manchon tout autour. Les étamines des magnolias se distinguent par l’absence de filament (la tige qui les porte) si bien qu’elles ne ressemblent pas du tout à l’image habituelle que l’on connaît, en trois parties distinctes ; les anthères contenant le pollen s’ouvrent vers l’intérieur. a ce moment-là, le pollen qu’elles contiennent est comme extrudé, rejeté si bien qu’il tombe au pied des étamines.
-

- Premier matin de l’ouverture d’une fleur : les étamines sont serrées autour de l’axe avec les pistils au-dessus

-

- Début d’après-midi du premier jour : les étamines commencent à s’écarter et à tomber ; les pistils au-dessus sont pleinement réceptifs

-

- Chute des étamines les plus à l’extérieur

Peu après l’ouverture, les étamines encore fermées commencent juste à s’écarter doucement ; dans les deux jours qui suivent, elles s’ouvrent et tombent tandis que celles plus à l’intérieur se déploient à leur tour vers l’extérieur. Ainsi, les étamines et leur pollen, écartées, s’accumulent en amas à la base des tépales en creux. Ce déballage désordonné de pollen et d’étamines sert de terrain de jeu et de repas pour des insectes qui s’y promènent dont des scarabées (Coléoptères). En se promenant, en s’accouplant et en se nourrissant, ils deviennent vite couverts de pollen (notamment les cétoines poilues) et peuvent ainsi transporter le pollen qui constitue bien la ressource nutritive primaire fournie par cette fleur.
-

- Le deuxième jour, les étamines s’accumulent à la base des pétales

-

- Vue de l’extérieur : on devine à l’intérieur de ce pétale un amas d’étamines tombées

Gynécée
Le gynécée (l’ensemble des nombreux pistils : de 40 à 50 en moyenne) se trouve concentré sur la massue terminale de l’axe central et forme comme une sorte de cône. Bien que très serrés, ils sont libres et non soudés. Chacun d’eux porte une sorte de bec crochu, le stigmate, la partie chargée de réceptionner le pollen pour assurer la fécondation. chaque pistil contient un ou deux ovules.
-

- Stigmates crochus réceptifs en début de floraison : la partie supérieure papilleuse capture le pollen

-

- Pistils dégagés en fin de floraison

Dès l’ouverture de la fleur, les stigmates sont réceptifs alors que les étamines n’ont pas commencé à s’ouvrir ; il y a donc un décalage dans le temps entre les deux sexes et on parle de protogynie puisque la fleur est d’abord femelle (pistils). Ce dispositif limite les échanges pollen/stigmates au sein de la fleur et donc l’autopollinisation défavorable pour la formation de graines viables. Quand les étamines libèrent leur pollen à partir du second jour, les stigmates tendent à ne plus être réceptifs. Ce mode de fonctionnement (femelle d’abord puis mâle) semble correspondre à la situation ancestrale originelle chez les plantes à fleurs ; la protandrie (mâle d’abord) bien plus répandue ne serait apparue que dans un second temps.

Au troisième jour, les étamines sont toutes tombées et les tépales amorcent leur déclin
Après la fanaison et la chute définitive des tépales desséchés, il ne restera plus que l’axe central avec sa base rouge portant les cicatrices des étamines et sa massue de pistils devenus chacun un fruit sec contenant une ou deux graines. D’abord verts à rosés, ces fruits vont se durcir et sécher avant de s’ouvrir selon une fente ventrale : on parle de follicules. Ces fruits et leurs graines feront l’objet d’une quatre chronique.
-

- Les strobiles se voeint à peine sur le fond du feuillage persistant dense.

-

- Ensemble de fruits (strobile) : chaque fruit est marqué par une fente ventrale

-

- Strobile mûr : les fruits s’ouvrent et on voit apparaître les graines rouge vif

Pollinisateurs
-

- Cétoine grise (Coléoptère) installée dans une fleur de magnolia à grandes fleurs

-

- Petit scarabée sur les stigmates d’une fleur de Magnolia hybride

Classiquement, dès la fin du 19ème siècle, à partir d’observations faites sur des magnolias plantés dans des parcs en Europe, on avait avancé que les coléoptères étaient les principaux pollinisateurs en accord avec l’idée que ce mode de pollinisation correspondait à celui des fleurs primitives. Mais, au moins chez nous, si on observe ces fleurs, on voit aussi de fréquentes visites d’abeilles domestiques et d’abeilles solitaires. Dans son milieu natal en Louisiane, des études ont montré qu’il y a certes des visites de coléoptères qui véhiculent du pollen (voir ci-dessus) mais aussi de fréquentes visites d’abeilles domestiques et d’au moins une espèce locale d’abeille solitaire ; or, le comportement de ces dernières montre une forte corrélation avec les transformations internes de la fleur sur les trois ou quatre jours de sa floraison. Ceci pointerait en faveur donc d’un rôle central des abeilles. Mais d’aucuns font remarquer que les abeilles visitent les fleurs surtout au stade « mâle » (étamines ouvertes) et participent en fait peu à la pollinisation effective. Cependant les stigmates élaborent des secrétions sucrées riches en hexose qui jouent un peu le rôle du nectar. Donc, même s’il y a encore controverse, il semble bien que les fleurs de nombre de magnolias ne soient pas ou plus que des fleurs à scarabées : il y a probablement eu une évolution récente dans une partie de la lignée vers une pollinisation par des hyménoptères (abeilles et bourdons).
-

- Abeille visitant une fleur en fin de floraison, largement ouverte

-

- Abeille au bord d’une fleur là aussi en fin de cycle (3ème jour)

Guides
D’autres faits convergent vers cette dernière hypothèse. La majorité des magnolias américains ont des fleurs blanches unies (le rouge est plus fréquent chez les asiatiques) sans véritables repères visuels pour guider les visiteurs. Mais si on éclaire leurs fleurs en lumière ultra-violette, on découvre qu’elles arborent alors des motifs contrastés susceptibles d’attirer les abeilles qui, elles, voient dans l’ultra-violet. Des photos prises dans la semi-obscurité et sous UV montrent que l’axe central apparaît alors rouge vif tandis qu’une fluorescence bleue souligne les tépales ; or, le bleu reste la couleur favorite des hyménoptères en général ! On a montré aussi que cette réflectance des UV évoluait dans la fleur pendant les quatre jours de floraison : d’abord forte sur les stigmates des pistils, elle passe ensuite sur les étamines, suivant ainsi l’évolution de la fleur (protogynie).
Les fleurs du magnolia à grandes fleurs disposent d’un autre argument de choc pour attirer ses pollinisateurs et qui flatte aussi notre odorat : un extraordinaire parfum capiteux, perceptible en s’approchant de la fleur en coupe. L’étude chimique montre sa richesse en une kyrielle de substances dont des terpènes tels que le géraniol, l’ocimène (qui se retrouve dans le basilic) ou le myrsène. La composition varie beaucoup d’une espèce à l’autre mais aussi d’un individu à l’autre avec sans doute des populations chimiques différentes. Par contre, la composition ne varie pratiquement pas pour un arbre donné pendant les quatre jours de la floraison ce qui pointerait vers l’attraction spécifique d’un seul type de pollinisateur.
On sait par ailleurs que les feuilles des magnolias endommagées par des attaques d’insectes libèrent et fabriquent par induction des substances chimiques volatiles du même type (terpènes) destinées à repousser les attaquants herbivores ou à attirer leurs parasites ou prédateurs. On pense donc que les voies métaboliques de synthèse de ces substances répulsives ont été transférées dans les fleurs et y ont subi des modifications (oxygénation des composés notamment) qui en fait des composés attractifs dans le cadre de la pollinisation ! L’évolution n’est décidément qu’un perpétuel recyclage !
BIBLIOGRAPHIE
- Comparative floral anatomy and ontogeny in Magnoliaceae. F. Xu and P. J. Rudall. Pl. Syst. Evol. 258: 1–15 (2006)
- Magnolia flower – the living crystal. Beata Zagórska Magnolia, 2011 – sophia.smith.edu
- The floral biology of Magnoliaceae. L. B. Thien et al. 1996 – pdfs.semanticscholar.org
- Floral scents, leaf volatiles and thermogenic flowers in Magnoliaceae. H. Azuma et al. Plant Species Biology (1999) 14, 121-127.
A retrouver dans nos ouvrages
Page(s) : 102-103

Page(s) : 20

