Ratites et Gondwana : une (trop) belle histoire !
Paleognathae

Autruche d’Afrique dans son milieu naturel au Kenya. Photo D. Bermudez
Le nom Ratites désigne l’ensemble des oiseaux « coureurs » qui ont perdu la capacité de voler tels que autruches, nandous, émeus, … Ce terme scientifique fut construit en 1816 sous la forme ratitae à partir du latin ratis pour radeau, allusion au sternum (os de la poitrine) plat dépourvu de crête osseuse ou bréchet ; il permettait de séparer ces oiseaux des autres, pourvus d’un bréchet et nommés carinates (pour carène). Passé dans la langue française en 1836 sous sa forme ratite, ce nom a quand même réussi à percer un peu dans le langage courant. Il s’agit en effet d’oiseaux spectaculaires tant par leur taille pour la majorité d’entre eux que par leur inaptitude à voler ; ils sont de plus presque toujours représentés jusque dans les plus petits zoos.
Evidemment, on a envie de savoir pourquoi ces grands oiseaux étranges ont perdu la capacité de voler et depuis quand. Cette question de l’histoire évolutive des Ratites anime la communauté scientifique depuis deux siècles et a fait l’objet de vifs débats. Vers les années 1980-2000, on était arrivé à un consensus quasi-général sur une histoire associée à celle du supercontinent Gondwana, une belle histoire que l’on trouve dans de nombreux ouvrages de référence ou de vulgarisation (2) et que nous relatons dans cette chronique. Mais, à l’aube du 21ème siècle, de nouvelles données moléculaires convergentes vont faire voler en éclats cette belle histoire (à laquelle j’ai adhéré bien sûr en son temps !) pour écrire une tout autre histoire (pas plus « laide ») que nous exposerons dans une seconde chronique.
Six petites familles
Passons rapidement en revue les ratites avec ses six familles (1).
-

- Autruche avec son long cou flexible

-

- Autruche au repos

-

- Chaque patte porte deux gros doigts

Les autruches ou Struthionidés comptent deux espèces africaines, l’une au cou et pattes rouges (autruche d’Afrique) et l’autre au cou et pattes grises foncées d’Afrique de l’Est (autruche de Somalie) ; leurs pattes puissantes pourvues de deux doigts seulement leur permettent d’atteindre des pointes de vitesse de 70 km/h.
-

- Nandous. Noter l’ouverture de l’oreille rarement visible chez la majorité des oiseaux !

-

- Nandou

-

- Le plumage est composé de plumes aux barbes très lâches, typique des Ratites

-

- Les pattes puissantes portent trois doigts

Les nandous ou Rhéidés (trois espèces) vivent dans les milieux ouverts sud-américains et possèdent des pattes avec trois doigts.
-

- Deux gravures représentant des casoars : un timbre ….

-

- .. et une gravure extraite d’un ouvrage d’Alcide d’Orbigny

-

- Les émeus sont de très proches parents des Casoars

-

- Les pattes portent aussi trois doigts mais sans les griffes redoutables du casoar.

La famille des Casuariidés regroupe d’un côté les trois espèces de Casoars des forêts tropicales denses d’Australie et de Nouvelle-Guinée et l’Emeu d’Australie. Les premiers se distinguent par la présence d’une sorte de casque osseux sur la tête et un doigt doté d’une redoutable griffe utilisée comme arme de défense.
-

- Deux espèces de Kiwis ou Aptéryx sur des timbres


Les Aptérygidés forment une famille vraiment différente avec cinq espèces toutes en Nouvelle-Zélande connues sous le nom de Kiwi ou Aptéryx. Ce sont des oiseaux de taille réduite (1 à 3 kg), avec un long bec fin dont les narines se trouvent tout au bout, des ailes très réduites, des plumes transformées en vibrisses à la base du bec et des pattes courtes avec des doigts aptes à fouir. Leur biologie présentent diverses originalités : des mœurs nocturnes, un régime alimentaire à base de vers, la capacité de creuser des terriers et la ponte d’un seul œuf « énorme », le plus gros connu chez les oiseaux par rapport à la taille du corps (25%) !
-

- Photo célèbre du paléontologue R. Owen auprès d’un squelette de Moa

-

- Moa géant sur un timbre néo-zélandais

-

- Reconstitution d’un oiseau-éléphant ou Aepyornis de Madagascar.

Restent deux familles d’oiseaux géants qui se sont éteintes au cours de la période historique et qui ont donc « croisé » les hommes … pour leur grand malheur ! Les moas ou dinornithidés vivaient en Nouvelle-Zélande avec neuf espèces (au moins) réparties en six genres ; les espèces les plus grandes atteignaient 3,6m de haut pour un poids d’environ 250 kg. Les derniers ont disparu autour de 1300 à 1400 avec la colonisation de l’archipel par les Maoris. L’Aepyornis ou oiseau-éléphant de Madagascar mesurait lui aussi jusqu’à 3,5m de haut mais pour un poids de 350 à 450 kg ce qui en fait l’oiseau le plus lourd ayant jamais existé. Il semble avoir disparu dès l’an 1000, victime sans doute de la colonisation humaine de la grande île. On retrouve des œufs à coquille encore intacte atteignant un volume de 9 litres !
On est frappé par la disparité de ce groupe et surtout par l’éclatement extrême de leur répartition ; la petite taille des familles indique clairement qu’il s’agit probablement des « miettes » restant d’un vaste groupe très ancien.
Des palais anciens
On a très tôt associé les Ratites avec un groupe d’oiseaux volants d’Amérique du sud, les Tinamous réunis dans un ordre à part les Tinamiiformes. Ces oiseaux ressemblent fortement, en apparence, à des pintades brunes ; ils se nourrissent au sol et sont capables de voler, sans être de bons voiliers, à la manière des perdrix avec leurs ailes courtes et arrondies. On en dénombre 48 espèces dans 9 genres, réparties dans les forêts tropicales et milieux plus ou moins ouverts d’Amérique du sud et centrale. Contrairement aux Ratites, ils possèdent un sternum avec un bréchet même si celui-ci est percé d’une profonde fenêtre latérale. On les associait aux Ratites car ils partagent avec eux une architecture osseuse du palais particulière qualifiée « d’ancienne » car plus proche de celle des Crocodiliens par exemple. On les réunit donc dans le groupe des Paléognathes (mot-à-mot : anciennes mâchoires) par opposition au reste des oiseaux actuels qui forment les Néognathes avec un palais différent. Les tinamous partagent par ailleurs un autre caractère biologique original avec les Ratites : c’est le mâle qui couve les œufs.
-

- Deux gravures anciennes représentant des tinamous


Les caractères anatomiques puis les données moléculaires accumulées à partir des années 1980 avaient conduit à un consensus sur trois points. Les tinamous sont le groupe-frère (le plus proche parent actuel) des Ratites. On peut les réunir dans le groupe des Paléognathes qui est le plus ancien groupe de la classification des Oiseaux actuels. Les Ratites forment de leur côté un groupe monophylétique, i.e. issu d’un ancêtre commun qui était avait perdu l’aptitude à voler ; autrement dit, l’aptérisme, la perte de cette capacité à voler, n’aurait eu lieu qu’une fois dans l’histoire du groupe des Ratites. Ce schéma va voler en éclats à l’aube du 21ème siècle (voir la seconde chronique sur les Ratites) et n’est désormais plus d’actualité.
Le tempo du Gondwana
Un des points qui intriguait le plus les scientifiques, c’est la répartition très éclatée des familles de Ratites (en y ajoutant les tinamous) sur les masses continentales de l’Hémisphère sud des six familles qui les composent : la carte de synthèse ci-jointe illustre cette répartition. Comment des oiseaux non volants avaient-ils pu se disperser à ce point sur des régions aussi éloignées et séparées par de vastes espaces océaniques ? Entre 1980 et 2000 on pensait avoir enfin élucidé ce mystère avec une théorie cohérente basée sur un parallèle entre la séparation des branches successives des Ratites et le démantèlement du supercontinent de Gondwana en autant de continents ou grandes îles.
Rappelons brièvement l’histoire du Gondwana : ce supercontinent s’était formé il y a environ 600 millions d’années et réunissait dans l’Hémisphère sud des masses continentales aujourd’hui séparées en une seule : Afrique, Amérique du sud, Inde, Antarctique, Australie. A partir de – 160 Ma, toujours dans le cadre de la tectonique des plaques, le Gondwana va commencer à se disloquer lentement avec la séparation de l’Inde et de l’Afrique ; l’Inde va ensuite complètement se détacher et entamer sa longue route vers l’Hémisphère nord ; vers – 80Ma, la Nouvelle-Zélande se détache puis ce sera l’Australie et la Nouvelle-Guinée, ….
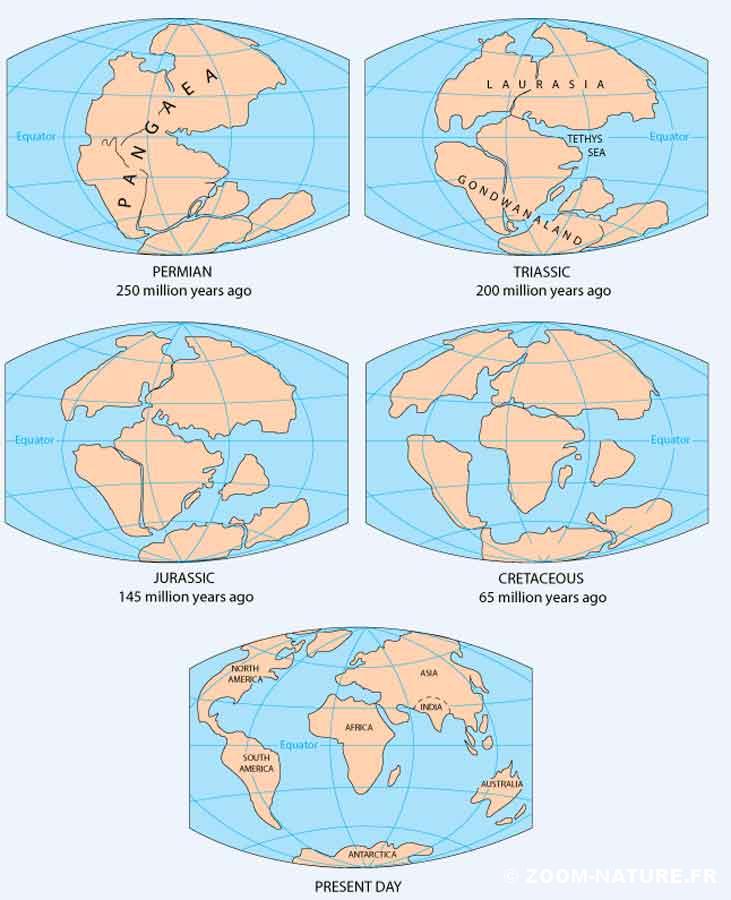
Cartes successives des mouvements des plaques avec la dislocation de l’ancien supercontinent Gondwana. D’après le site http://pubs.usgs.gov
L’idée était donc que l’évolution des Ratites, supposés tous aptères non volants, avait en quelque sorte suivi ces étapes successives.
La vicariance biogéographique
On appelle ainsi le phénomène qui conduit à la séparation d’une population ancestrale (ici, l’ancêtre commun des ratites réparti sur la masse gondwanienne) en plusieurs populations séparées du fait de la séparation des masses continentales. Ainsi, par exemple, la séparation de l’île de Madagascar du continent africain aurait conduit à l’isolement de populations ancestrales qui auraient ensuite évolué en isolement vers les Aepyornis ; et ainsi de suite. Ainsi se trouvait contourné l’obstacle de la dispersion impossible même si on invoquait aussi dans les détails la possibilité de dispersion vers des îles via des radeaux flottants de végétation comme cela est connu pour d’autres groupes non volants. Au passage, il est drôle de voir à quel point cette image du radeau a collé aux Ratites : depuis le sternum plat en « radeau » (voir l’étymologie), en passant par ces radeaux de végétation et sans oublier la vieille image des continents-radeaux dérivant les uns par aux autres chère à Wegener !
Cependant, en dépit de son côté très séduisant, cette théorie qui avait fini par s’imposer et être vulgarisée, présentait quelques écueils majeurs. Les reconstitutions phylogénétiques de l’histoire des six familles des Ratites ne collaient pas parfaitement avec l’ordre de succession de séparations des masses continentales ; il fallait recourir à des hypothèses secondaires pour faire coller les deux. Ainsi, les données moléculaires pointaient les moas comme non apparentés directement aux kiwis (Aptérigydés) ce qui impliquait deux colonisations indépendantes de la Nouvelle-Zélande, particulièrement isolée dans le Pacifique.
L’obstacle des fossiles
L’autre écueil qui a émergé concerne les fossiles progressivement découverts. La plupart des formes non volantes sont apparues au-delà de la limite Cretacé/Tertiaire donc nettement après la grande dislocation du Gondwana déjà alors bien avancée. Ainsi, on trouve très peu de fossiles de Paléognathes au début du Tertiaire alors que selon l’hypothèse du Gondwana, ces grands oiseaux auraient déjà dû être bien présents et avec de fortes probabilités d’être fossilisés vu leur taille. Plus encore, la découverte de paléognathes fossiles dans l’Hémisphère nord impliquerait la dispersion de ces derniers dès le Crétacé. En plus certains d’entre eux étaient non volants (comme Palaeotis trouvé en Europe ; début Eocène) et d’autres volants comme les Lithornis d’Europe et d’Amérique du nord (début du Tertiaire). Tous les Paléognathes se sont ensuite éteints au cours du Tertiaire dans l’Hémisphère nord avant le retour des autruches en Afrique. La divergence de ces dernières des autres ratites avait dès les années 2000 été située comme nettement postérieure à la séparation de l’Afrique du reste du Gondwana.
Bref, il y avait déjà dans cette pourtant belle histoire des signes de craquements, annonciateurs de son « explosion en plein vol » ( !) à l’aube du 21ème siècle ; ceci fera l’objet de la seconde chronique sur les Ratites avec la nouvelle version de leur histoire revisitée.
BIBLIOGRAPHIE
- Bird families of the World. D. W. Winkler et al. Lynx Ed. 2015
- The ancestor‘s tale. R. Dawkins. Weidenfield et Nicholson. 2004
- Tinamous and Moa Flock Together: Mitochondrial Genome Sequence Analysis Reveals Independent Losses of Flight among Ratites. MATTHEW J. PHILLIPS, GILLIAN C. GIBB, ELIZABETH A. CRIMP, AND DAVID PENNY. Syst. Biol. 59(1):90–107, 2010
- Phylogenomic evidence for multiple losses of flight in ratite birds. J. Harsham et al. 13462–13467 ; PNAS ; 2008
- Ratite Nonmonophyly: Independent Evidence from 40 Novel Loci. JORDAN V. SMITH, EDWARD L. BRAUN, AND REBECCA T. KIMBALL. Syst. Biol. 62(1):35–49, 2013
A retrouver dans nos ouvrages
Page(s) : 416-418

Page(s) : 517-533

