Ratites : une autre histoire
Dans la chronique Ratites et Gondwana, nous avons présenté les Ratites, ce groupe d’oiseaux qui réunit autruches, émeu, casoars, nandous, kiwis, moas et aepyornis, tous aptères i.e. inaptes au vol même s’ils ont conservé leurs ailes. Ils forment avec le groupe des Tinamous sud-américains, des oiseaux volants, un groupe ancien, les Paléognathes, à la base de l’arbre de parentés des Oiseaux actuels. Nous avons aussi présenté la théorie qui prévalait jusqu’à très récemment quant à leurs origines : un ancêtre commun non volant unique à partir duquel les six familles actuelles ou récemment éteintes se seraient diversifiées en suivant la dislocation du supercontinent Gondwana dans l’Hémisphère sud ; ainsi se seraient isolées les unes après les autres des lignées différentes sur les masses continentales devenues isolées du fait de la dérive des continents. Nous avons terminé cette chronique en précisant que cette histoire qui faisait autorité est désormais obsolète avec au moins cinq études parues depuis 2008 (3, 4, 5 et 6) qui, chacune avec des données moléculaires différentes, aboutissent à la même conclusion : les Ratites tels qu’ils sont définis ne forment pas un groupe avec un ancêtre commun ! Ceci change complètement la donne et conduit à reconsidérer entièrement l’histoire de ces oiseaux : tel est l’objet de cette seconde chronique qui va donc « démolir » la précédente et témoigner de l’évolution permanente des connaissances scientifiques.
Les ratites ne peuvent plus être conservés dans leur ancienne version compte tenu du positionnement des Tinamous.
Les autruches à la base
Les comparaisons récentes de séquences de gènes nucléaires différents ou de gènes mitochondriaux confirment toutes la position basale des autruches (Struthionidés) au sein des Paléognathes, i.e. qu’elles constituent la lignée la plus ancienne de tout l’ensemble Ratites et Tinamous. Le recours à l’horloge moléculaire permet même de situer la divergence de cette première branche autour de – 100 à – 90Ma, soit bien après le début de fractionnement du Gondwana qui avait commencé dès – 160Ma avec la séparation de l’Afrique. D’emblée, cette donnée remet en doute l’hypothèse de la vicariance évoquée dans l’autre chronique.
Cette position basale donc ancienne rejoint une découverte récente surprenante à propos du sommeil des autruches (1) : la part de sommeil paradoxal est la plus élevée de tous les oiseaux et durant celui-ci s’intercalent des phases brèves de sommeil lent ; autrement dit, il n’y a qu’un type de sommeil mixte et pas deux types biens séparés dans le temps. Or, on ne retrouve une telle situation que chez les Echidnés et les Ornithorynques, dans le groupe des Monotrèmes, la branche la plus ancienne des Mammifères !
Mais alors d’où viennent les autruches ? Les fossiles les plus anciens connus de cette lignée (Paleotis) proviennent d’Europe et datent de l’Eocène. Ensuite, on ne trouve trace des autruches en Afrique que tardivement à partir du Miocène en même temps qu’avaient lieu de grands échanges de faunes entre l’Eurasie et l’Afrique avec la fermeture de la Méditerranée. Il y aurait donc eu soit un événement de dispersion vers le nord très tôt depuis le Gondwana soit même une différenciation sur place, dans l’Hémisphère nord, de la première lignée. Les autruches n’auraient donc pas une origine gondwanienne du « grand Sud » et seraient nées « out of Africa »!

L’Autruche d’Afrique a vraiment un air « reptilien » et elle n’est pas sans rappeler les dinosaures du genre Struthiomimus ! Photo D. Bermudez
La place des tinamous
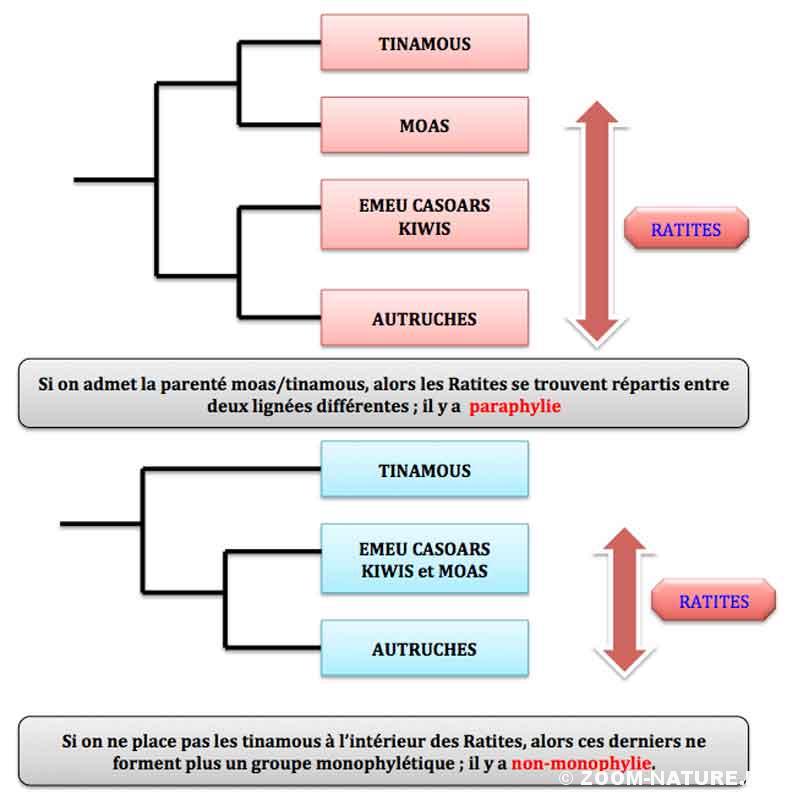
L’autre convergence inattendue de ces études moléculaires concerne le positionnement des Tinamous : ils ne sont pas à l’extérieur du groupe des Ratites comme on le supposait jusqu’alors mais au cœur de celui-ci et après les autruches ; autrement dit, on aboutit à une situation où en plein milieu de l’arbre qui réunit des oiseaux non volants vient s’insérer une lignée d’oiseaux volants ! Donc, les Ratites ne sont plus un groupe monophylétique, issu d’un ancêtre commun puisqu’il faut leur adjoindre une autre lignée. En toute rigueur, on ne devrait plus utiliser ce terme dès lors qu’on se situe dans une perspective de classification (tout comme les groupes des Rapaces par exemple : voir la chronique Les Rapaces ne sont plus ce qu’ils étaient) mais compte tenu de l’usage courant de ce terme, on continue à l’employer par commodité. Les scientifiques ont donc défini un nouvel ordre qui inclut les ex-Ratites et les tinamous : les Struthioniformes, en référence à la famille basale des autruches.
-

- Deux gravures anciennes représentant des tinamous


En dépit de l’avancée sur les données moléculaires, on n’arrive pas encore à être certain de la place exacte des Tinamous au sein des « non-autruches » actuels. Deux hypothèses ressortent néanmoins : le groupe frère des Tinamous pourrait être soit l’ensemble des Ratites australasiens actuels (Emeu, Casoars et Kiwis) clairement parents entre eux, soit les Nandous. La seconde hypothèse semble la plus évidente a priori compte tenu de la proximité géographique (nandous et tinamous sont sud-américains actuellement) ; mais si on intègre dans les données la famille éteinte des Moas de Nouvelle-Zélande, on obtient un très fort indice d’une étroite parenté Tinamous-Moas ! Ce qui veut dire aussi que l’histoire des Moas de Nouvelle-Zélande ne peut être associée à celle des autres membres locaux du groupe (Kiwis, Casoars et Emeu).

Un des arbres de parentés le plus probables entre les Paléognathes (d’après 5)
Plusieurs pertes de la capacité à voler
Comment expliquer la présence d’oiseaux volants au cœur d’un groupe aptère ? On pourrait invoquer que l’ancêtre commun était bien aptère et que la faculté de voler soit réapparue dans la lignée des tinamous. Mais ce scénario semble très peu probable au regard de ce que l’on observe actuellement : dans près de 18 familles actuelles, on trouve des espèces aptères ; rien que dans la famille des Rallidés, on estime que la capacité de voler a été perdue plus de 100 fois indépendamment sur de nombreuses îles du Pacifique ! Il est donc très facile de perdre cette faculté d’autant qu’on oublie que voler requiert de fortes dépenses en énergie et un investissement important dans la masse musculaire ; chaque fois que le contexte le permet (niches écologiques sans prédateurs par exemple), l’évolution vers l’aptérisme se manifeste. La tendance concomitante à évoluer vers des tailles de plus en plus grandes dans cette lignée des Paléognathes constitue un facteur favorisant supplémentaire. A l’inverse réacquérir l’aptitude au vol serait quasiment impossible car près de 20Ma séparent la branche des autruches de celle de tinamous.
Ceci signifie que l’ancêtre commun à tous ces oiseaux était volant et que l’aptérisme, la perte de la capacité à voler, n’a pas émergé une seule fois à l’origine mais plusieurs fois : une fois dans la première lignée des autruches et au moins deux autres fois dans la lignée des nandous et dans celle des Ratites australasiens. L’autre conséquence importante c’est que l’on peut désormais faire appel à des évènements de dispersion par le vol pour expliquer l’installation de nouvelles lignées sur des îles ou continents séparés depuis un moment !
Les données fossiles confirment cette nouvelle vision : on trouve des paléognathes aussi bien volants que aptères et ce, jusque dans l’Hémisphère nord. Un autre fait surprenant c’est que les lignées de type Lithornis, des paléognathes volants répandus au début du Tertiaire, se sont éteintes de manière indépendante en Eurasie, en Amérique du nord, à Madagascar, en Nouvelle-Zélande, en Australie et en Antarctique à l’époque encore libre de glaces. Seule la lignée des tinamous a survécu en Amérique du sud.
Un nouveau scénario de dispersion

Cartes des mouvements des continents (source : 7) au Secondaire et Tertiaire
Le cadre général de la nouvelle histoire se situe donc après la crise de fin du Crétacé (et non plus bien avant) et la disparition des Dinosaures non aviens et d’une partie des mammifères ; la diminution temporaire relative de la pression de prédation (avant l’explosion des mammifères) aurait libéré de nouvelles niches propices à la perte de la capacité à voler à partir d’ancêtres volants tout en augmentant de taille. La lignée des Lithornis fossiles, oiseaux paléognathes volants, commence un peu avant la crise Crétacé et les premiers grands paléognathes aptères apparaissent quelques millions d’années après cette limite comme en Argentine.
Les parentés se clarifient entre familles actuelles et éteintes. Ainsi, l’oiseau-éléphant ou Aepyornis de Madagascar n’est pas, comme on le croyait, parent proche des autruches mais appartiendrait à la lignée australasienne Casoars-Kiwis-Emeu. La séparation de Madagascar du reste des masses continentales s’est faite vers – 110Ma et une dispersion par vol de l’ancêtre a pu se faire plus tard alors que Madagascar était encore « à portée » d’ailes !
Deux scénarios de dispersion à partir de l’Amérique du sud vers l’Australie (longtemps associé à l’Antarctique) et la Nouvelle-Zélande peuvent être avancés : soit en une seule fois avec ensuite un retour vers l’Amérique du sud des tinamous, soit, plus probablement, en deux fois, une pour les ancêtres des kiwis-casoars-émeu et une autre fois pour l’ancêtre des Moas toujours à partir des Tinamous. Il faut garder à l’esprit que même si les masses continentales s’éloignaient de plus en plus, leurs distances respectives sont restées un long moment « abordables » pour des traversées en vol, sans parler de l’existence possible d’archipels intermédiaires aujourd’hui disparus.
De fortes convergences
Si malgré toutes ces divergences répétées, les « Ratites » se ressemblent en grande partie, c’est qu’il y a eu une forte convergence en fait liée à la perte du vol et à l’adoption d’un mode de locomotion de type course et la tendance vers de grandes tailles.
Il y a quand même un cas très à part, celui des Kiwis ou Aptérygidés de « toute petite taille » (relativement aux autres !) et très différents à bien d’autres égards (voir leur présentation dans la chronique sur Ratites et Gondwana). Jusqu’à récemment, on ne disposait pas de fossiles anciens pour comprendre leur origine. On vient de trouver en Nouvelle-Zélande, leur patrie, (2) des fossiles d’un Aptérygidé daté de – 19Ma ; la reconstitution dresse le portrait d’un oiseau volant de petite taille ! Ceci confirme donc l’arrivée de ces oiseaux sur l’archipel par vol au-dessus de l’océan et indépendamment des ancêtres qui ont donné la lignée des Moas. On prétendait jusqu’à peu que les kiwis résultaient d’une évolution par nanification à partir d’ancêtres de grande taille (et on pensait alors aux moas) à cause notamment de leurs très gros œufs qui leur permettent d’avoir des poussins très nidifuges, capables de quitter le nid très tôt.

Les Kiwis sont vraiment à part !
Il reste donc encore beaucoup à découvrir au sujet des Ratites et de leur histoire décidément mouvementée. D’aucuns pourraient se lamenter que l’ancienne histoire liée au Gondwana soit ainsi balayée en si peu de temps mais la nouvelle version vaut largement l’ancienne ! Pour peu que notre point de vue émotionnel d’humains ait une quelconque importance sur de tels sujets !
BIBLIOGRAPHIE
- Ostriches Sleep like Platypuses. Lesku JA, Meyer LCR, Fuller A, Maloney SK, Dell’Omo G, et al. PLoS ONE 6(8) (2011)
- Miocene fossils show that kiwi (Apteryx, Apterygidae) are probably not phyletic dwarves. T. H. Worthy et al. Paleornithological Research 2013. Proceed. 8th Internat. Meeting Society of Avian Paleontology and Evolution
- Ratite Nonmonophyly: Independent Evidence from 40 Novel Loci. JORDAN V. SMITH, EDWARD L. BRAUN, AND REBECCA T. KIMBALL. Syst. Biol. 62(1):35–49, 2013
- Phylogenomic evidence for multiple losses of flight in ratite birds. J. Harsham et al. 13462–13467 ; PNAS ; 2008 vol. 105 no. 36
- Tinamous and Moa Flock Together: Mitochondrial Genome Sequence Analysis Reveals Independent Losses of Flight among Ratites. MATTHEW J. PHILLIPS, GILLIAN C. GIBB, ELIZABETH A. CRIMP, AND DAVID PENNY. Syst. Biol. 59(1):90–107, 2010
- Genomic Support for a Moa–Tinamou Clade and Adaptive Morphological Convergence in Flightless Ratites. Allan J. Baker Oliver Haddrath, John D. McPherson, and Alison Cloutier. Mol. Biol. Evol. 31(7):1686–1696. 2014
- Carte de la Pangée et du Gondwana tirée du site : http://pubs.usgs.gov/gip/dynamic/historical.html
A retrouver dans nos ouvrages
Page(s) : 416-418

