Les couleurs vues à travers les yeux des oiseaux

09/08/2022 Jusque dans les années 1970, la communauté scientifique a considéré que les oiseaux voyaient les couleurs des objets naturels de leur environnement (dont leurs congénères) de la même manière que les humains : on croyait, avec toute la « modestie évolutive » qui caractérise l’espèce humaine, que les couleurs que nous percevons étant une réalité objective valaient pour tous les vertébrés. Pourtant, dès les années 1960, on savait que divers insectes dont les abeilles ne percevaient pas les couleurs comme nous puisqu’elles étaient sensibles aux ultra-violets (UV) que notre œil ne perçoit pas. Néanmoins, on ne chercha pas à savoir si cette possibilité concernait d’autres groupes dont les oiseaux. Depuis les années 1970, on a découvert qu’en fait presque tous les oiseaux possédaient cette faculté et que leur sensibilité aux couleurs était considérablement plus étendue et précise que la nôtre. Cette nouvelle vision du monde sensoriel des oiseaux a conduit à reconsidérer complètement la manière dont ils perçoivent leur environnement mais aussi la signification et la fonction des couleurs innombrables qu’ils affichent sur leurs plumages.
Nous allons donc ici exposer brièvement et simplement comment fonctionne la vision des couleurs chez les oiseaux. Pour autant, cette connaissance scientifique ne nous permettra pas de savoir comment les couleurs de leur environnement leur apparaissent réellement : eux seuls le savent et nos cerveaux sont incapables de l’imaginer faute d’avoir les mêmes équipements sensoriels et les capacités associées de traitement des informations.
Et la couleur fut …
Jusqu’au 17ème siècle, on considérait que la couleur d’un objet était une propriété inhérente à celui-ci. Or, dès qu’on place un objet dans l’obscurité il « perd » cette couleur à nos yeux. En fait, elle est indissociable de la lumière ambiante qui éclaire l’objet observé : sans lumière, pas de couleur. Or, la lumière naturelle « n’a pas de couleur » ; on parle de lumière blanche pour la désigner. C’est I. Newton qui révéla ce lien couleur/lumière en interprétant la dispersion de la lumière blanche à travers un prisme : on fait ainsi apparaître les sept composantes principales colorées visibles par notre œil sous forme de bandes colorées selon un continuum graduel, le spectre visible. On peut le retrouver par la formule mnémotechnique VIBVJOR (violet, indigo, bleu, vert, jaune, orange, rouge). En pratique, il existe tous les intermédiaires entre ces différentes couleurs. On observe exactement le même phénomène avec les arcs-en-ciel où les rideaux de gouttelettes d’eau décomposent la lumière incidente.

La physique explique que la lumière est un type d’onde électromagnétique (au même titre que les ondes radio) qui se compose d’une infinité de longueurs d’ondes différentes (exprimées en nm ou nanomètres, des milliardièmes de m, soit des millionièmes de mm). A chaque longueur d’onde lumineuse correspond une couleur différente : par exemple autour de 380nm, nous sommes dans le bleu (toujours à nos yeux). Quand nous regardons un « objet », nous percevons sa couleur à travers les ondes lumineuses qu’il nous renvoie et qui frappent nos yeux. L’œil humain ne capte que les ondes comprises en gros entre 400 et 700nm (le spectre visible humain) ; nous ne percevons donc pas les ondes qui sont juste en-dessous des 400nm (les ultra-violets) ni juste au-dessus de 700nm (les infra-rouges), même si notre peau peut ressentir la chaleur qu’elles génèrent.
Vision des couleurs
C’est plus fort que nous : nous parlons toujours d’abord de nous, humains, et je vais poursuivre encore dans ce sens ; malgré tout, c’est avec ce qu’on connaît le mieux, nous-mêmes (a priori), qu’on peut espérer comprendre « les autres » pas trop distants de nous, les oiseaux étant des vertébrés tétrapodes comme nous.
On peut définir la vision des couleurs comme la capacité à discriminer des objets éclairés via les longueurs d’ondes lumineuses qu’ils renvoient. Le siège de cette capacité se situe au niveau de la rétine, cette membrane fine reliée directement au cerveau par le nerf optique. Elle possède des cellules photoréceptrices spécialisées, les cônes (à cause de leur forme), qui renferment un pigment visuel : une grosse molécule protéique colorée (opsine) liée avec une molécule de vitamine A ; chaque cône est couplé avec une cellule nerveuse ou neurone.
Quand un photon (particule élémentaire de lumière) frappe une molécule de pigment, celle-ci l’absorbe ce qui change sa forme et déclenche une chaîne de réactions biochimiques qui aboutissent à moduler la quantité de médiateur chimique libéré par le cône. Ce médiateur passé dans le neurone va y générer un influx nerveux électrique transmis au cerveau et analysé.
Pour qu’une couleur soit perçue, il faut que le cerveau puisse comparer les influx issus d’au moins deux types de cônes de sensibilité différente, i.e. réagissant à des longueurs d’onde (donc des couleurs) différentes. Certains scientifiques ajoutent à cette définition de la vision des couleurs la capacité de les apprendre et de s’en faire une représentation interne permettant de ressentir la sensation colorée. C’est en cela que la couleur est une construction perceptuelle et non pas une réalité intangible.
L’espèce humaine a une rétine dotée de trois types de cônes qui diffèrent par le pigment visuel qu’ils renferment ; chaque type réagit (i.e. engendre des influx dans le neurone) quand il est touché par des longueurs d’onde comprises dans une bande étroite avec un pic de sensibilité maximale (sensibilité spectrale). Ainsi, on distingue les cônes « bleus » sensibles autour de 430nm : on les qualifie de cônes S (pour short, ondes courtes) ; les cônes « verts » (ou M pour moyen) réagissent autour de 530nm et les cônes « rouges » (L pour longs) autour de 560nm. Leurs bandes de sensibilités se chevauchent un peu : ainsi les cônes verts et rouges, assez proches, perçoivent de nombreuses couleurs communes. On dit que l’espèce humaine a une vision trichromatique (3 types de cônes) ce qui correspond aux 3 couleurs primaires (bleu, vert, rouge) ; les autres couleurs sont composées par superposition (voir le RVB des écrans). En fait, c’est bien notre cerveau qui reconstitue toutes les couleurs en traitant les millions d’informations qui lui parviennent simultanément.

Ajoutons que par contre, la sensation visuelle de luminosité (la luminance) ne requiert qu’un seul type de photorécepteur dans la rétine, les bâtonnets ; nous nous en servons pour apprécier la forme des objets et leur texture tout particulièrement dans l’obscurité.
En 4 couleurs
Bon, assez parlé de nous ; désormais, place aux oiseaux, le vrai sujet de cette chronique. Néanmoins, ce passage par l’Homme nous aura doté du bagage élémentaire indispensable pour comprendre la suite. L’essentiel de ce qui vient d’être exposé au niveau du fonctionnement global vaut tout autant pour les oiseaux que pour d’ailleurs la majorité des vertébrés : on y retrouve une rétine, plusieurs types de cônes avec des sensibilités spectrales différentes, des bâtonnets, des opsines comme pigments visuels, le traitement neurologique de la perception, …
Mais derrière cette apparente unité se cachent des divergences profondes qui rendent la vision des couleurs des oiseaux radicalement différente de la nôtre.
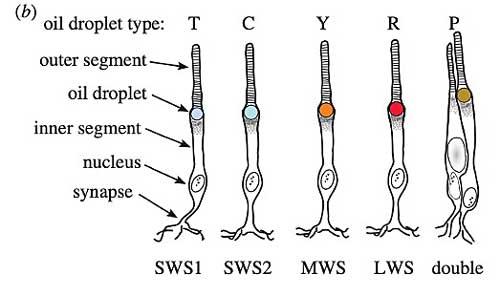
Types de gouttelettes (en haut) : T/transparent ; C/clair ; Y/jaune ; R/rouge
Types de cônes de gauche à droite : UVS ; VS ; Médian (vert) ; Long (Rouge) ; cône double
L’écrasante majorité des oiseaux possède 4 types de cônes dans sa rétine et non pas trois (voir ci-dessus). On retrouve les cônes rouges L (543-571nm), les verts (497-510nm) et les bleus (430-463nm) ; mais s’y s’ajoute un quatrième type : les cônes sensibles aux violets (cônes VS) (soit des couleurs à la limite de la perception humaine) et/ou les cônes sensibles aux UV (cônes UVS) qui captent globalement entre 362 et 426 nm. Donc les oiseaux disposent d’un spectre visible nettement plus étendu que le nôtre en allant en deçà des 300nm dans l’ultra-violet. Les oiseaux ne sont donc pas trichromatiques mais tétra (ou quadri) chromatiques.
On trouve par ailleurs dans la rétine des cônes doubles formés par un cône principal et un cône accessoire appariés intimement par deux ; ils serviraient dans la détection des mouvements, la reconnaissance de motifs et la texture de la vision.
Voir les UV
De ce fait nous ne pouvons pas même imaginer les couleurs que voient les oiseaux dès lors qu’elles incorporent des UV réfléchis par des objets. Or, on sait que de nombreux objets naturels (dont les plumages des oiseaux) renvoient les UV (contenus dans la lumière blanche) et que donc ils sont perçus différemment par les oiseaux. On pense que, devant une même scène, un oiseau dispose d’une vision bien plus subtile et doit repérer plein de détails que nous ne voyons pas.
La tétrachromatie des oiseaux ne surprend pas les biologistes car on la connaît chez divers vertébrés : des poissons (comme le poisson-zèbre), des amphibiens et des reptiles ; les analyses phylogénétiques démontrent que ce caractère devait exister dès l’ancêtre commun de tous les vertébrés ; il a ensuite évolué différemment selon les lignées successives ou été perdu, notamment chez les mammifères.

Les Ratites dont l’autruche forment la lignéela plus basale des oiseaux actuels ; ils n’ont pas de cônes UVS …. 
… sauf chez les Nandous sud-américains
Chez les oiseaux, la sensibilité UV n’est pratiquement pas présente dans les lignées basales (sauf chez les nandous au sein des ratites) ; ainsi les Galliformes et les ansériformes (voir la chronique) possèdent des cônes Violets Sensibles VS mais pas de cônes UVS. Tout indique que l’ancêtre commun des oiseaux était tétrachromate avec des cônes VS qui fonctionnent avec une opsine particulière (SW1). Ensuite, dans les lignées dérivées, à cinq reprises au moins de manière indépendante, des pigments sensibles aux UV ont évolué à partir du pigment ancestral VS par des mécanismes inconnus chez tous les autres vertébrés. Autrement dit, la sensibilité UV des oiseaux reste bien à part au sein des vertébrés de part son origine. On la trouve ainsi chez les perroquets et leur groupe frère, l’immense groupe des Passereaux (avec néanmoins des exceptions ponctuelles), chez les mouettes et goélands ou chez des oiseaux sud-américains aux plumages vivement colorés, les trogons.

Deux groupes d’oiseaux ont secondairement perdu la tétrachromatie : les chouettes et hiboux ou strigiformes (voir la chronique) et les manchots (Sphénisciformes) ; dans les deux cas, on peut supposer que cette perte est à relier avec la vie dans des « environnements visuels » très particuliers : la nuit ou sous l’eau.
La petite gouttelette qui change tout
Les cônes des oiseaux possèdent en plus une autre particularité originale : sous le segment externe qui capte la lumière, au sommet du segment interne se trouve une gouttelette huileuse sphérique colorée qui occupe tout le diamètre de ce segment. Ce caractère est connu chez d’autres vertébrés mais les oiseaux (ainsi que les tortues) sont les seuls à avoir de telles gouttelettes vivement colorées en rouge (cônes L), orange ou jaune vif (cônes M).
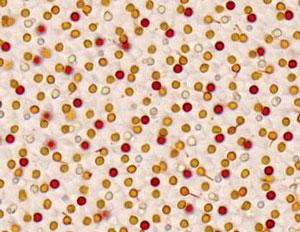
Rétine d’oiseau au microscope avec les gouttelettes colorées des cônes
Une partie de la lumière qui a traversé le pigment situé au-dessus (segment externe) réussit à atteindre la gouttelette en dessous. Celle-ci, du fait de sa forme et de son contenu très concentré en pigments, agit comme une microlentille très puissante qui coupe certaines longueurs d’onde et agit comme un filtre qui rétrécit au final la bande de longueurs d’onde qui ressortent ; ainsi, elles réduisent le chevauchement entre les cônes de sensibilité proche. Autrement dit, elles améliorent l’acuité de perception de la couleur. Cet effet vient se surajouter à la perception primaire en l’améliorant considérablement.
La coloration des gouttelettes dans des tons jaune, rouge et orange indique la présence de caroténoïdes (voir la chronique), ces pigments colorés que les oiseaux ne peuvent synthétiser directement et qu’ils récupèrent (soit directement, soit en les resynthétisant) à partir de leurs aliments. Or, ces pigments se retrouvent par ailleurs dans de nombreux plumages où ils servent, entre autres, de signaux visuels de communication (voir la chronique). Le type et la concentration des caroténoïdes dans chaque gouttelette déterminent sa capacité à transmettre la lumière et à la modifier au passage.
Chez la poule, chaque gouttelette renferme un mélange complexe de caroténoïdes avec une variété dominante qui détermine l’effet sur la lumière. Les cônes L, M et S renferment respectivement de l’astaxanthine, de la zéaxanthine et de la galloxanthine. Cette palette de caroténoïdes permet donc d’affiner considérablement le filtrage tonal des cônes et la vision des couleurs. Les cônes UV et V sensibles sont quant à eux dotés d’une gouttelette claire ou transparente (type T) ainsi que les cônes doubles.
Outre cet effet, on a avancé de manière hypothétique d’autres fonctions indirectes de ces gouttelettes : protéger des UV délétères ; améliorer la capture des photons lumineux ; détecter le champ magnétique terrestre.
Folie évolutive
L’évolution réitérée de la sensibilité UV (voir ci-dessus) de manière indépendante dans diverses lignées suppose qu’à chaque fois des mécanismes de transformation du pigment concerné ont été sensiblement différents. On se trouve donc devant un processus multiforme.
On ne fait que commencer à explorer en détail, groupe par groupe, la présence ou pas de cette capacité et ses modalités. Et des surprises sont au rendez-vous notamment dans l’immense groupe des passereaux qui a connu une explosion évolutive récente. Ainsi, on a découvert que chez le bobolink ou goglu des prés, un passereau de la famille des Ictéridés, très connu en Amérique que les cônes bleus (S) étaient absents ; cela signifie qu’il faudrait en fait étudier en détail, espèce par espèce la structure de la rétine, un travail dantesque en perspective. Encore plus surprenante cette découverte chez des moucherolles (genre Empidonax ; famille des Tyrranidés), oiseaux insectivores qui chassent les oiseaux en vol : la gouttelette huileuse unique de certains cônes est remplacée par un complexe de mégamitochondries surmontées de centaines de microgouttelettes orange. Ces cônes différents sont répartis çà et là dans la rétine surtout dans la région centrale. Autrement dit, il y a probablement bien d’autres dispositifs encore inconnus à découvrir au niveau des cônes des oiseaux ; et il reste aussi à comprendre la fonction précise de telles innovations.


On ne fait aussi que commencer à comprendre les conséquences de cette forme de vision dans la vie des oiseaux qui en sont dotés. Ainsi, on a démontré que les faucons crécerelles perçoivent les pistes d’urine laissées par les petits rongeurs sous forme de « trainées UV réfléchissantes » qu’ils détectent depuis le ciel ; même cachés par l’herbe les rongeurs restent repérables. On sait aussi que certains fruits sur un décor de feuillage environnant sont hautement détectables par les oiseaux frugivores via les UV renvoyés. Et il reste le vaste domaine des parades nuptiales où des taches colorées du plumage prennent encore plus d’importance du fait de la détection des UV réfléchis.
Il est clair que la vision a été soumise à une très forte pression évolutive du fait de l’acquisition du vol chez les oiseaux ce qui rend l’usage de ce sens encore plus crucial. Ce n’est par hasard si les yeux des oiseaux occupent un volume considérable dans la boîte crânienne.

Et encore, ici nous n’avons envisagé que la partie de la vision qui concerne la perception des couleurs ; dans d’autres domaines comme l’acuité, la capacité de fixation, la rapidité, la vision binoculaire, … les oiseaux montrent des capacités sidérantes. Nous étions bien présomptueux de les rabaisser à notre mode de vision qui paraît, par contraste, comme étant peut-être un des moins performants des vertébrés amniotes. Mais ne tombons pas dans ce piège de la hiérarchie et considérons les oiseaux pour ce qu’ils sont : des êtres qui ne peuvent que nous émerveiller et aussi nous faire rêver en pensant à ce que nous pourrions percevoir si nous avions leurs yeux.
Bibliographie
The Visual Ecology of Avian Photoreceptors Nathan S. Hart Progress in Retinal and Eye Research Vol. 20, No. 5, pp. 675 to 703, 2001
Through birds’ eyes: insights into avian sensory ecology Graham R. Martin J Ornithol (2012) 153 (Suppl 1):S23–S48
Avian Color Vision and Coloration: Multidisciplinary Evolutionary Biology Andrew T. D. Bennett and Marc Thery vol. 169, supplement the american naturalist january 2007
A complex carotenoid palette tunes avian colour vision Toomey MB, Collins AM, Frederiksen R, Cornwall MC, Timlin JA, Corbo JC. 2015. J. R. Soc. Interface 12: 20150563.
A novel cellular structure in the retina of insectivorous birds. Luke p. tyrrell et al. Scientific RepoRtS (2019) 9:15230
