Balsamine de l’Himalaya : une invasion à la dérive
Impatiens glandulifera

Depuis son introduction en Europe à partir des années 1840, la balsamine de l’Himalaya est passée du statut de belle ornementale des jardins d’eau à celui d’invasive honnie dont on cherche à se débarrasser à tout prix. En un peu plus d’un siècle, elle a envahi tous les grands bassins fluviaux et colonisé les forêts qui longent les cours d’eau, formant des peuplements denses et fournis. Un tel dynamisme ne manque pas d’interpeller et au-delà des particularités écologiques de cette espèce qui lui confèrent des avantages certains, il faut interroger son mode de dispersion pour comprendre une telle expansion toujours en cours. Comme les nombreuses autres espèces du genre balsamine ou impatiente (Impatiens), elle possède des fruits explosifs capables de projeter les graines jusqu’à cinq mètres autour (voir la chronique consacrée aux aspects balistiques de ce mode de dispersion). Mais cela n’explique pas comment elle a pu coloniser des milliers de kilomètres de bords de rivières. Evidemment, on pense à la possibilité logique d’un transport par le courant des rivières (hydrochorie) : or, curieusement, en dépit de nombreuses études, cette possibilité n’avait jamais été explorée ; on invoquait plutôt la dispersion par les activités humaines (anthropochorie) comme moteur de dispersion à longue distance. Une étude anglaise vient d’étudier cette hypothèse de l’hydrochorie et d’apporter enfin des éléments de réponse tangibles (1).

Les rivières sont des corridors naturels.
Possible ?
La majorité des pieds de balsamine ne sont pas juste au bord de la rivière : autrement dit la projection explosive des graines même jusqu’à cinq mètres (voir la chronique) ne suffit pas pour atteindre le courant ! L’hypothèse d’un transport par l’eau a longtemps été négligée car, après leur projection hors des capsules, les graines flottent peu ou très mal !

Les crues changent la donne en apportant d’une part du courant et d’autre part en s’étalant hors du lit principal. (rivière la Sioule près de Pontgibaud)
Oui mais, lors de crues avec un très fort courant, elles se trouvent néanmoins entraînées, mélangées avec les sédiments charriés par les flots et redéposées bien en aval dans un lit de vase idéal pour la germination de cette espèce annuelle qui apprécie les sols humides enrichis en matières nutritives. D’autre part, des observations en laboratoire et sur le terrain montrent que les fruits entiers non éclatés (pas complètement mûrs) peuvent, eux, flotter plusieurs jours et même dans une eau calme. De même des fragments (valves) de fruits éclatés mais portant encore des graines accrochées (voir la chronique sur les fruits explosifs) peuvent aussi flotter au fil de l’eau.
-

- Manchons de débris déposés par une crue en marge de la rivière, témoins de la hauteur de la lame d’eau
-

- Cette banquette alluviale a été parcourue par une crue comme en témoignent les herbes sèches couchées
-

- Les crues s’étalent souvent dans la forêt alluviale en bordure, le milieu d’élection de la balsamine; les graines transportées sont déposées en même temps que les alluvions (sable, vase) et les débris charriés.
Donc, potentiellement au moins, cette hypothèse d’un transport à longue distance par l’eau (hydrochorie) tient a priori la route mais dans le cadre d’évènements de crues ou inondations seulement sans entrer avec en contradiction avec l’existence de l’autre moyen de dispersion explosif qui fonctionne à courte distance.
-

- Fruit encore vert ouvert pour montrer les graines

-

- Au cnetre, une germination de balsamine sur un lit de vase.

Pister les graines

Si l’hypothèse d’un transport des fruits et/ou graines par les rivières est juste, alors elle doit impacter les échanges génétiques entre populations installées ou en voie d’installation au long des rivières dans le sens du courant. Pour en vérifier la réalité, les chercheurs britanniques ont sélectionné deux bassins fluviaux, l’un en Irlande et l’autre au Pays de Galles, colonisés par des balsamines et avec des bifurcations, i.e. des affluents et des confluences. Ils ont échantillonné les populations présentes au long de ces rivières connectées ou pas entre elles en recherchant dans l’ADN de ces plantes des marqueurs génétiques particuliers, des groupes de gènes partageant une même fonction.
-

- Le sens du courant va imposer la direction de la dispersion par hydrochorie

-

- Ces marques nous rappellent la hauteur de certaines crues au bord de l’Allier : un sacré potentiel de transport !

Les résultats montrent clairement une augmentation de la diversité génétique de ces marqueurs « au fil de l’eau » quand on descend vers l’aval avec l’arrivée successive d’affluents : ceci suggère fortement une dispersion par l’eau dans le sens du courant. Un exemple illustre bien ce processus : une population située sur un affluent est génétiquement très différente d’une autre située pourtant à moins d’un kilomètre mais sur une rivière non connectée (bassin versant différent) alors qu’elle est bien plus apparentée à une autre située à huit kilomètres en aval sur la même rivière !
Deux faits biologiques peuvent expliquer cette tendance : les échanges de pollen (côté « mâle ») et/ou les déplacements de graines (côté « femelle » maternel). Or, pour le pollen, les principaux pollinisateurs sont ici les abeilles domestiques dont le comportement reste fortement contraint par la nécessité de retrouver à la ruche ce qui limite d’autant l’étendue des flux de pollen. Donc, ce serait bien par voyage des graines que la diversité génétique se propage d’amont en aval.
De l’importance du débit
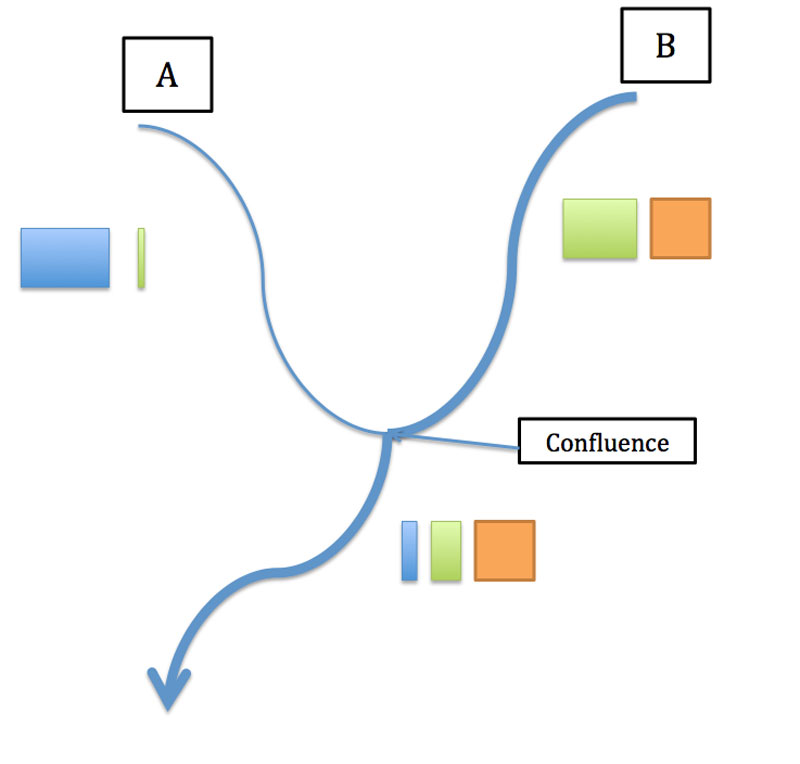
Schématisation de la situation décrite ci-dessous (d’après 1) ; les carrés colorés représentent les trois grands groupes de gènes analysés ; la taille des carrés dépend de leur représentation dans chacune des populations
Un autre exemple révélé dans cette étude illustre en plus l’importance de la structure du réseau hydrologique sur le processus. En amont de la confluence de deux rivières (que nous appellerons A et B), on constate que les populations de la rivière A partagent un groupe de gènes particulier (« bleu ») tandis que celles de la rivière B en partagent deux autres différents (« vert et orange »). Une population échantillonnée à une petite centaine de mètres de la confluence en aval renferme les trois groupes mais avec une forte prédominance des « vert et orange ». Comment expliquer une telle répartition inégale des groupes de gènes qui devrait logiquement être plus équilibrée ? La rivière B est beaucoup importante que la A et en période de crues son courant domine largement : autrement dit, le cours d’eau le plus puissant en débit lors des crues impose son tempo et son flux de graines ! La population de la rivière A tirerait son originalité (un groupe de gènes propre) de ses origines : elle est issue d’une population semée dans un jardin (donc implantée par l’homme) il y a 22 ans près de la source de la rivière ; elle est donc issue d’une introduction unique (venue d’ailleurs) et a évolué « en vase clos » et n’a pas envoyé de graines (ou très peu) au delà de la confluence.
-

- La balsamine d el’Himalaya est une ornementale appréciée dans les jardins humides pour la rapidité de sa croissance et sa floraison estivale ; ici, dans un parc floral

Vous aurez remarqué que pour que ce processus fonctionne il doit y avoir une « source » initiale en amont qui ensuite se propage donc vers l’aval en parcourant potentiellement de grandes distances en peu de temps.
Et l’homme dans tout çà ?
C’est là que l’homme, vecteur initial, intervient. Par exemple, un jardinier sème des balsamines de l’Himalaya comme ornementales dans son jardin humide ; si la population semée prospère (et notamment si le jardin est près d’un cours d’eau qui entretient une forte humidité) elle peut se répandre de proche en proche alentours (par voie explosive) et atteindre l’eau libre et ensuite se propager vers l’aval. On parle d’événement fondateur.
-

- Les bords des grandes rivières sont devenus des dépotoirs à ciel ouvert où l’on dépose sans scrupule toutes sortes de déchets

-

- Les déchets de jardin (ici au bord de la rivière Allier) sont d’importants agents de propagation de plantes exotiques cultivées.

La liste potentielle de tels évènements est longue :
– travaux d’entretien des berges qui envoient des fragments de plantes dans l’eau et font exploser les capsules à tour de bras
– transfert par des matériels ou des véhicules : par les pneus, sur les machines d’entretien, les engins agricoles, …
– déplacements du bétail avec des graines collées sous les sabots
– déblais de terre issue de travaux de terrassement près de maisons déposés au bord de l’eau.
-

- Les tas de déblais, le plus souvent illégaux, se multiplient dans ces milieux riverains peu fréquentés (pas vu, pas pris !)

-

- Les déblais issus de travaux urbains véhiculent des graines de plantes cultivées comme cette rose trémière.

Pour autant, cette étude prouve qu’en dépit de leur multiplicité potentielle, ces évènements fondateurs liés aux activités humaines restent assez limités (la plante ne se naturalise souvent pas à moyen terme) mais, par contre, une seule population installée suffit ensuite à alimenter le processus de descente aval à grande échelle et dans un temps relativement limité.
Contrôle et avenir
Comme la balsamine pose souvent des problèmes écologiques dans les milieux qu’elle colonise et semble même favoriser les processus d’érosion des berges par sa présence, on cherche à mettre en place des actions cohérentes pour contrôler son expansion. De cette étude, il ressort que les interventions pour l’éradiquer devraient cibler en priorité les populations les plus en amont sur une rivière donnée sans oublier les affluents s’ils sont assez puissants pour apporter leur flux de graines.
-

- Massif fleuri en plein développement : en cas de crue tardive, ce peuplement sera détruit vu la fragilité des tiges de cette plante

-

- La balsamine est une annuelle dont les peuplements disparaissent entièrement en hiver (à part les tiges sèches !)

L’expansion de la balsamine semble donc étroitement associée aux crues et inondations, lesquels devraient connaître une certaine augmentation dans les décennies à venir du fait du changement climatique. Mais ce n’est pas si simple (2) car il faut prendre en compte le moment où ont lieu les crues. En effet, les crues en fin de printemps (comme celles de juin 2016 en France) s’avèrent dévastatrices pour les balsamines qui sont des plantes annuelles peu enracinées aux grandes tiges fragiles. L’impact négatif maximal se situe pour une crue ayant lieu juste avant le début de maturation des fruits : les populations peuvent être anéanties sans que des fruits mûrs soient emportés.
Pour autant, il reste toujours quelques individus qui échappent et vont produire des graines en grand nombre et très rapidement la progression reprend même si elle a subi un coup d’arrêt temporaire. Restera à voir comment tout cela évoluera si des crues tardives de fin de printemps se multiplient ? cela limitera peut-être la balsamine mais à quel prix pour les reste de la végétation et de la faune en pleine période de reproduction !

Vers la rivière !
BIBLIOGRAPHIE
- Genetic evidence for predominantly hydrochoric gene flow in the invasive riparian plant Impatiens glandulifera (Himalayan balsam). Heather M. Love et al. Annals of Botany 112: 1743–1750, 2013
- Floods affect the abundance of invasive Impatiens glandulifera and its spread from river corridors. Jan Cuda et al. Diversity and Distributions, (Diversity Distrib.) (2017) 23, 342–354
A retrouver dans nos ouvrages
Page(s) : 288-89

