Rouilles : cinq vies entre deux hôtes

29/04/2021 Dans le monde des champignons, foisonnant et souvent étrange à nos yeux, certains groupes n’ont pas du tout bonne presse. Déjà, si vous n’êtes pas un champignon comestible, vous avez d’emblée perdu l’essentiel des chances que l’on s’intéresse à vous. Si en plus vous êtes parasite et microscopique, responsable de maladies sur des plantes cultivées, alors on ne s’intéressera à vous que sous l’angle de l’attaque et de l’éradication. Vous n’êtes qu’un nuisible, un dégénéré, une erreur de la nature qui n’a d’intérêt que quand on réussit à l’éliminer. Tel est le cas des rouilles, un groupe de champignons parasites de végétaux vivants qui se manifestent par des taches ou pustules colorées sur les feuilles et les tiges, honnis du grand public. Et pourtant, nombre d’entre elles offrent des scènes hautes en couleur et, surtout, dès que l’on s’intéresse à elles pour en savoir plus sur leur mode de vie, s’ouvre un monde sidérant avec sa galerie d’adaptations remarquables et des facultés hors du commun. Bienvenue donc chez les rouilles : laissez vos préjugés en berne et ouvrez les yeux pour goûter à ce monde étonnant qui a beaucoup à nous apprendre.

Alternance
Le groupe des rouilles, les Pucciniales des scientifiques, réunit au moins 7800 espèces de champignons microscopiques, réparties dans 170 genres à travers le monde. Ce nom de rouille (repris en anglais: rust) fait allusion à la couleur brun-rouge à orangé de certaines de leurs spores, les « cellules reproductrices » susceptibles d’engendrer par germination des filaments, base du mycélium ou appareil végétatif du champignon : lors de leur libération, ces spores donnent souvent aux plantes parasitées un aspect « rouillé » typique et bien connu.
Toutes les rouilles se comportent en parasites se nourrissant des tissus de végétaux vivants (biotrophiques) sans les tuer, tout au moins dans le court terme. Elles peuvent infecter (terme utilisé à leur égard compte tenu des pathologies qu’elles engendrent) la plupart des groupes de plantes dites vasculaires : les fougères, les conifères et les plantes à fleurs. Le cycle de vie annuel d’une grande partie d’entre elles se singularise par une alternance entre deux espèces de plantes hôtes très différentes. On distingue l’hôte dit principal sur lequel elles peuvent survivre indéfiniment sous des conditions favorables et sur lequel elles effectuent la part la plus « active » de leur reproduction et l’hôte dit alternatif sur lequel elles ne restent que peu de temps. Néanmoins, le passage par cet hôte alternatif n’a rien de secondaire puisque pour beaucoup d’espèces il est obligatoire pour que le cycle reparte sur l’hôte principal l’année suivante.

Epine-vinette commune dans une haie 
Feuilles dentées et tiges épineuses
La seconde originalité de cette dualité concerne la nature des espèces hôtes : il s’agit d’espèces non apparentées, souvent très éloignées dans la classification végétale. Ainsi, la rouille du blé (Puccinia graminis) alterne son cycle entre les céréales comme hôte principal et … un arbuste sauvage, l’épine-vinette commune comme hôte alternatif ! Une de ses très proches cousines, la rouille brune du blé (P. triticina) alterne elle entre les feuilles du blé et une espèce de plante herbacée rare, le pigamon jaune. La rouille dite « treillis du poirier » alterne entre les feuilles des poiriers et les pousses de certaines espèces de … genévriers, soit un grand écart entre une angiosperme d’un côté et une gymnosperme de l’autre ! Cette vie « entre deux » soulève d’emblée des questions évolutives passionnantes : comment réussissent-elles à infecter et parasiter deux hôtes sans parenté en utilisant des processus d’infection différents sur les deux ? Comment survivent-elles en cas d’absence d’un des deux hôtes dans l’environnement ?
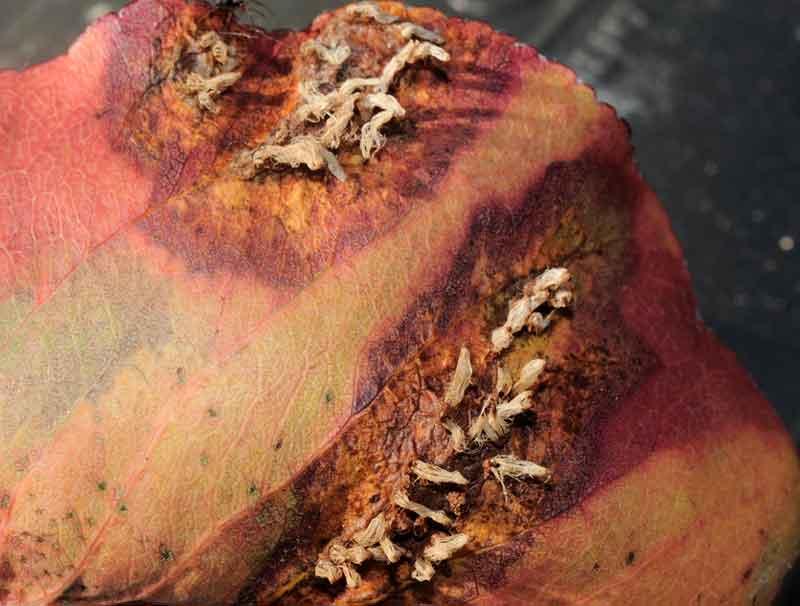
Gymnospsorangium fuscus sur feuille de poirier : surnommée le treillis du poirier 
Ecies de Gymnosporangium claviiforme sur une feuille d’aubépine 
Télies sèches de G. claviiforme sur un genévrier commun
Premier hôte
Non contentes d’alterner entre deux hôtes sur un cycle annuel, les rouilles s’y développent en passant par une succession de cinq stades ou formes de vie différentes, chacune élaborant des structures qui produisent et libèrent des spores différentes. Ces structures se forment sur le mycélium, un ensemble de filaments ou hyphes, implantés dans les tissus des végétaux infectés et insinués entre les cellules, qui constituent « le corps » du champignon. Nous allons prendre l’exemple ultra-classique, et sans doute le plus étudié pour des raisons économiques, de la rouille noire des céréales en commençant par les deux stades sur l’hôte alternatif, l’épine-vinette. Pour s’y retrouver, on numérote les cinq stades et spores associées de 0 à IV.
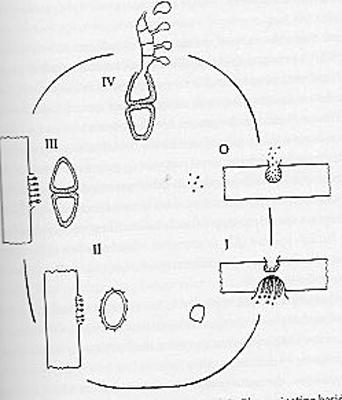
L’infection des feuilles de l’épine-vinette commence sur la face supérieure avec l’apparition de pustules en forme de mini-flacons faisant des taches en surface : ce sont les spermogonies (autrefois appelées pycnies), soit le stade 0 ; elles laissent sortir des paquets de filaments allongés en forme de vers annelés dont l’extrémité libère de microscopiques éléments unicellulaires, des spermaties (spores 0). Pour passer au stade suivant, ces spermaties issus d’une rouille donnée doivent entrer en contact avec les filaments émergeant d’une rouille de « polarité » différente : chez les champignons, on ne parle pas de sexe mâle ou femelle mais de polarité sexuelle + ou – avec des cellules sexuelles rigoureusement identiques (sauf au niveau de leur contenu génétique). Elles doivent donc être transportées vers d’autres spermogonies le plus souvent sur une autre épine-vinette (voir la dispersion). Quand une spermatie atteint ainsi un filament « opposé », son noyau y entre et fusionne avec celui du filament (fécondation).
Ce filament « fécondé » va se développer vers la face inférieure de la feuille et engendrer de nouveaux organes : des écidies ou écidioles (du mot grec oikidion, petite maison) ; c’est le stade I. Elles produisent, après une division réductrice ou méiose, des spores spécifiques ou écidiospores Ces écidies sont très visibles sous forme de pustules en coupe qui éclatent en libérant les spores I. Celles-ci sont incapables de germer sur l’épine-vinette et doivent donc atterrir sur une céréale, l’hôte principal pour que le cycle se poursuive.
Deuxième hôte
Arrivées sur une céréale, les spores I germent et donnent un nouveau mycélium qui pénètre dans les tissus et produit de nouveaux organes, des urédies (stade II) sous les feuilles ou sur les tiges ; elles s’ouvrent rapidement comme des fentes et libèrent des multitudes de spores de type II (urédospores) portées sur de longs pédoncules fins. Celles-ci vont infecter les plantes proches et propager très rapidement le parasite : on peut parler de phase épidémique. Une fois atterries sur une nouvelle plante (ou juste à côté), ces spores I germent, donnent un nouveau mycélium qui engendre de nouveaux organes (nette impression de se répéter !), des télies (stade III ; de telos, sommet achèvement) qui produisent des téliospores. Celles-ci, aux parois épaisses, ne se dispersent pas et sont capables d’hiberner dans les feuilles et tiges mortes ou dans le sol. Ainsi, en cours d’été, les urédies sont progressivement remplacées par des télies surtout sur les gaines des feuilles et des tiges où elles sont fermement accrochées. Elles jouent donc le rôle de stade de résistance pour passer l’hiver.
Le printemps venu, les téliospores germent sur les feuilles mortes et élaborent des organes allongés : des basides (stade IV) qui produisent des basidiospores. Ce nom parle tout de suite aux mycologues même simples amateurs éclairés car on le retrouve dans le nom du « grand » groupe, les Basdiomycètes (mycète = champigon) qui inclut la majorité de ceux que le grand public nomme spontanément des « champignons », i.e. aussi bien les agarics, les coulemelles, les tricholomes, les bolets, les amanites, … Effectivement, les rouilles font bien partie de ce vaste ensemble, même si leur mode de vie a conduit à leur extrême transformation.
Les basidiospores, une fois dispersées ne peuvent germer qu’à la surface d’une feuille d’épine-vinette sur laquelle elles vont générer le stade 0 : et c’est reparti ! Comment un cycle aussi compliqué avec autant de stades différents et à chaque fois des spores différentes avec chacune leur propre mode de dispersion a-t-il réussi à évoluer ? On pressent au vu du cycle que certaines étapes sont apparues en lien avec les cycles de vie respectifs des deux hôtes et leur calendrier de développement annuel : un hôte alternatif vivace versus un hôte principal annuel dont le feuillage ne se développe vraiment qu’à partir du milieu du printemps. Mais ceci ne constitue qu’une micro-explication face aux innombrables contraintes auxquelles sont confrontées les rouilles dans leur cycle.

Puccinia poarum sur Tussilage ou pas d’âne 
Détail des écies sous une feuille de pas d’âne 
Puccinia poarum sur le second hôte, une graminée (Pâturin, Poa) 
Urédies et télies sur feuille de pâturin
Le cycle ainsi exposé, d’une complexité maximale, ne concerne pas toutes les rouilles. Chez diverses espèces ou genres, on observe l’abandon de l’hôte alternatif : le cycle ne se déroule plus que sur un seul hôte. Ainsi la rouille des mauves (Puccinia malvacearum) se cantonne sur les mauves et autres genres apparentés dont les roses trémières (voir la chronique) dont les feuilles sont rapidement infestées au printemps ; on parle d’espèce autoécique (versus hétéroécique pour deux hôtes en alternance). Plus souvent, on observe une réduction du nombre de stades avec en premier lieu les urédies (stade II) qui disparaissent comme chez la rouille dite treillis du poirier (voir ci-dessus). Tout ceci montre l’extrême adaptabilité des rouilles notamment leur capacité à contourner par exemple la disparition d’un des hôtes.

Dessus de feuille de rose-trémière infectée 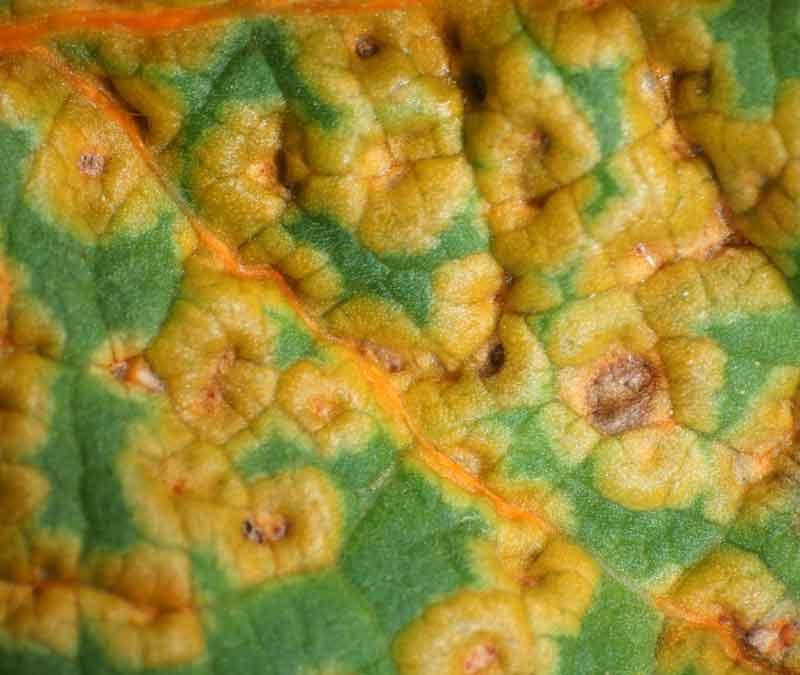
Taches laissées par les télies 
Télies vues par dessous (loupe) 
Dispersion
Pour quatre des cinq stades de ce cycle, des spores sont libérées et doivent être dispersées pour infecter soit le second hôte soit de nouvelles parties sur l’espèce infectée. Or, selon les stades, on a découvert que des mécanismes différents de dispersion sont mis en jeu, dont certains très élaborés.

Au printemps, les touffes émergentes de Falcaire (ombellifère) sont presque toutes infectées 
Feuilles infectées par P. sii-falcariae (hôte unique) 
Ecies mûres sous une feuille de falcaire
Le vent reste l’agent principal de dispersion compte tenu du caractère microscopique des spores comme chez la majorité des champignons. C’est le cas notamment pour les spores II (urédospores) qui colonisent massivement les plantes autour du pied infecté : ces spores de contamination ont une paroi très épaisse, pigmentée et épineuse et peuvent survivre en l’air plusieurs semaines ce qui leur laisse la possibilité de voyager sur de très grandes distances. En plus, les filaments qui les produisent sont allongés et placent ainsi les spores au-dessus de la surface de la feuille infectée, facilitant la prise en charge par le moindre courant d’air.
Pour les spores IV (basidiospores), intervient un mécanisme actif spectaculaire connu chez la majorité des champignons du groupe des Basidiomycètes (voir ci-dessus) : la dispersion par catapultage. Peu avant leur libération, deux gouttes de liquide rondes se forment, l’une vers la base et l’autre au sommet ; elles s’agrandissent et finissent par se réunir ce qui déclenche instantanément une « explosion » qui catapulte la spore. Cette projection résulte d’un processus dit de tension de surface qui nécessite une forte humidité atmosphérique : ainsi, cette dispersion a lieu le plus souvent de nuit au pic d’humidité. Inversement, s’il y a de l’eau libre au contact des spores, le processus ne peut avoir lieu : ceci explique la forme en chapeau ou en parapluie de la majorité des champignons « classiques » qui abrite les lamelles sur lesquelles sont fabriquées ces spores.

Mais le mode de dispersion le plus surprenant concerne les spermaties, les spores libérées par les spermogonies (stade 0). Prenons l’exemple de la rouille du cirse des champs (P. punctiformis), une espèce très répandue à cycle réduit sur ce seul hôte. Au printemps, les jeunes plants infectés de cirse des champs qui émergent se repèrent de loin à leur aspect jaunâtre et à l’odeur sucrée surprenante qui en émane ! Les feuilles sont couvertes de spermogonies qui libèrent des substances aromatiques volatiles telles que le benzaldéhyde (présent dans les pêches, framboises, fraises, ..), le 2-phényléthanol indole (odeur de rose avec notes de jacinthe) et le phénylacétaldéhyde (odeur de jacinthe). Ces parfums floraux attirent les insectes qui visitent ainsi les feuilles infectées et participent au transfert des spores (spermaties). Chez l’épine-vinette (voir ci-dessus), les spermogonies libèrent un liquide sucré équivalent du nectar : les pollinisateurs visitent ces organes et transportent ainsi les spermaties qui doivent rencontrer des filaments du « sexe » opposé pour que le cycle se poursuive. Il y a encore mieux chez certaines rouilles qui transforment complètement l’aspect des tiges infectées avec la formation de « pseudofleurs », des paquets de feuilles simulant une fleur et qui produisent du nectar sucré. L’exemple le plus connu est celui de la rouille du pois (Uromyces pisi) qui parasite les euphorbes petit-cyprès et à laquelle nous consacrerons une chronique entière.

Spermogonies jaune-orange et écies brunes sous les feuilles d’un cirse 
Urédies ou télies mûres
Infection
Pour la continuité du cycle, l’autre étape cruciale concerne la capacité des spores, une fois dispersées, à germer et à infecter la plante sur laquelle elles ont atterri avec le challenge du changement d’espèce d’hôte.

Les spores IV (basidiospores) pénètrent dans l’hôte directement via la cuticule, le revêtement de l’épiderme, sans développer de structure particulière. Les autres spores (et notamment les spores II de contamination) par contre développent au bout des filaments émis après germination une structure, un appressorium, une cellule spécialisée aplatie qui appuie et s’enfonce dans une cellule de l’hôte utilisant la pression de turgescence pour s’enfoncer ; ce dispositif peut même percer des films miroir sans tain réfléchissants ! C’est l’équivalent des suçoirs des plantes parasites comme les cuscutes (voir les chroniques).
Dans un premier temps, les spores doivent se fixer à la surface de la plante ; elles se collent à condition d’être hydratées et élaborent un coussinet d’adhésion ; une fois bien ancrées, elles libèrent des enzymes qui modifient les propriétés de la surface de la cuticule de l’hôte et cimentent le coussinet à la surface. La spore peut alors émettre son tube de germination qui se colle lui aussi à la surface. Pour que le fameux organe « perceur » (appressorium) se forme, il faut que le tube entre en contact avec un rebord infime d’au moins un-demi micron de hauteur. Cette réaction unique parmi les champignons relève d’une réponse automatique à une sensation tactile (thigmotropisme). Dès que le signal est perçu, l’organe perceur se développe en moins d’une heure. Sur les feuilles, la surface qui déclenche ce « réflexe » est la lèvre d’un stomate, l’une des innombrables micro-ouvertures qui parsèment la surface des feuilles et servent à échanger les gaz avec l’air (respiration et photosynthèse). Le tube de germination franchit l’ouverture et juste derrière les cellules de garde du stomate se renfle en une vésicule à partir de laquelle vont pousser de nombreux filaments qui s’insinuent entre et dans les cellules de la plante hôte via leurs « têtes perceuses ».
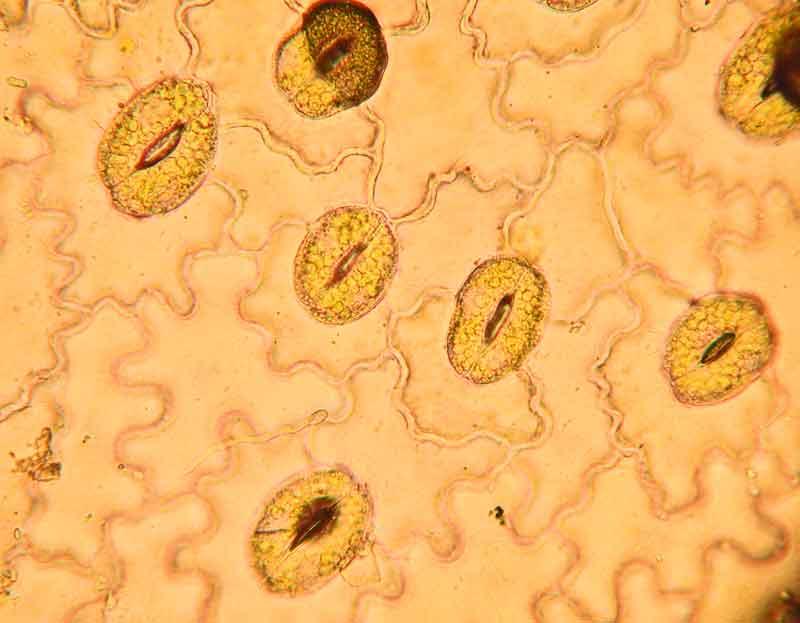
On reste abasourdi devant la précision et l’efficacité de ces mécanismes et surtout le fait qu’ils soient capables de s’adapter à des hôtes très différents qui disposent en général de leur coté de mécanismes de défense chimiques ou physiques différents. On remarque aussi la convergence de forme avec les plantes à fleurs parasites dans ces structures de pénétration de cellules.
Histoire(s)

Télies sur feuilles de buis (P. buxi) 
Télies vues par en dessous
Ce n’est pas par hasard si on connaît autant de détails sur le cycle de vie et les structures développées par les rouilles : un certain nombre d’espèces parasitent des plantes cultivées dont les céréales et peuvent causer des dégâts considérables avec des répercussions économiques et sociales qui accompagnent l’homme depuis la domestication des céréales. Elles sont donc un enjeu de recherche majeur en agronomie. Dans les années 1960, en pleine guerre froide, les Américains étudiaient même la possibilité de l’utiliser comme arme biologique !
On pense que les rouilles des céréales descendent des rouilles qui vivaient sur les graminées ancestrales et que les hôtes alternatifs (comme l’épine-vinette pour la rouille noire) se trouvaient dans les mêmes régions. La migration vers l’Ouest depuis le berceau du grand Moyen-Orient aurait commencé très tôt dans l’histoire de l’agriculture des céréales puisque dès – 3300 ans on en trouve des traces archéologiques en Israël.
La rouille noire des céréales a longtemps été « la bête noire » des agriculteurs et le lien avec la présence nécessaire de l’épine-vinette dans l’environnement proche a été suspecté il y a plusieurs siècles. Dès 1660 près de Rouen ou 1755 dans le Massachussetts, on a initié des campagnes d’éradication des épine-vinette près des champs de céréales. En 1918, après deux grandes épidémies en 1904 et 1916 qui avaient détruit 50% des récoltes de céréales dans l’Ouest américain, une campagne massive d’éradication fut lancée et qui s’est poursuivie à grande échelle jusque dans les années 30 et, localement, encore pendant plusieurs décennies. Globalement, ces campagnes ont permis d’atténuer la sévérité des épidémies de rouille : comme les spores II ont une durée de vie limitée et sont produites en quantité limitée, le rayon d’action potentiel d’un pied d’épine-vinette se limite à 3-4 kms environ. On a aussi retardé ainsi l’apparition de nouvelles races ou variétés en atténuant considérablement la phase de reproduction sexuée où se produisent les recombinaisons génétiques.
La démonstration expérimentale du cycle sur deux hôtes fut faite en 1864 par A. de Bary ; jusqu’alors on considérait que les stades sur l’épine-vinette correspondaient à une espèce à part nommée Aecidium berberidis. En fait, cette rouille peut se développer sur pas moins de 365 graminées hôtes différentes appartenant à 54 genres différents ; en pratique, on constate qu’il existe de nombreuses formes spécialisées adaptées à seulement une ou quelques espèces très proches ; ainsi, on connaît la rouille noire du blé, celle d’avoine ou celle de l’orge et elles s’hybrident entre elles ! La mise sur le marché de nouvelles variétés de céréales résistantes via notamment des hybridations avec des variétés anciennes a permis de limiter la portée de cette maladie.
Dans la Rome antique, dans la vaste galerie des dieux, figurait, Robigus, dédié à la protection des cultures contre les maladies et notamment contre la rouille des blés ; cette maladie facilement reconnaissable aux pustules rouge brique entre les nervures des feuilles a dû être associée à ce dieu infernal via la couleur rouge évoquant le feu. On célébrait ce dieu protecteur des cultures le 25 avril : cette date coïncide avec le début du développement de la rouille sur les céréales en pleine croissance en région méditerranéenne. La fête associée se nommait Robigalia au cours de laquelle on sacrifiait un chien (symbole du côté obscur) et un mouton (symbole de fertilité).
Cette fête a été reprise dans la religion chrétienne sous la forme des rogations, ces prières faites le 25 avril (fête de St Marc) et durant les trois jours qui précèdent l’Ascension pour attirer la bénédiction divine sur les récoltes et les travaux des champs. Anatole France en 1908 dans son ouvrage Jeanne d’Arc écrivait : « La veille de l’Ascension, aux Rogations ou Petites Litanies, les croix étaient portées par les champs ».

Une rouille sur des feuilles de pâquerette (P. obscura) 
Rouille spécifique (télies noires) de la moschatelline (Puccinia adoxae)
Bibliographie
Introduction to fungi. Third edition. J.Webster ; R.Weber. Cambridge University Press 2007
