La dispersion du merisier : la voie terrestre
01/12/2021 Dans les régions tempérées, le merisier sauvage colonise toute une gamme de milieux allant des forêts aux espaces agricoles ouverts sous forme d’arbres isolés en passant par les haies ou les bosquets. Pour se disperser, i.e. s’installer dans de nouveaux sites, il s’appuie sur ses fruits charnus (les merises) succulents dotés d’un noyau dur (drupe : voir la chronique sur les fruits charnus des Rosacées), consommés par des animaux au régime au moins en partie frugivore : la pulpe est digérée mais le noyau, particulièrement dur et résistant, se retrouve généralement intact dans les excréments après le transit intestinal. Ce processus qui concerne jusqu’à 60% des espèces d’arbres dans les forêts tempérées s’appelle l’endozoochorie (voir la chronique consacrée à ce mode de dispersion) et permet souvent des évènements de dispersion à longue distance compte tenu du temps parfois long nécessaire à la digestion complète et au rejet des excréments pour des animaux qui se déplacent avec de grands rayons d’action. Dans nos pays tempérés, ce processus a été relativement étudié dans sa version avienne, i.e. avec des oiseaux frugivores comme agents de dispersion, mais très peu ou pas du tout dans sa version « terrestre » avec comme agents des mammifères. De plus, la majorité des études concernent souvent des milieux forestiers ; mais qu’en est-il dans des environnements profondément perturbés par les activités humaines comme les vastes zones d’agriculture intensive ? Une équipe allemande a donc relevé le défi d’étudier cette dispersion des noyaux de merises par divers mammifères terrestres à travers toute une gamme de paysages allant de milieux forestiers à des zones agricoles intensives très simplifiées en passant par des zones agricoles extensives avec un certain nombre d’éléments semi-naturels (haies, bosquets, arbres isolés, prairies, …).

Voyages digestifs
Dans la vie des plantes à fleurs, la dispersion (ou dissémination) occupe une place centrale souvent méconnue du grand public car moins facile à percevoir que par exemple la pollinisation par les insectes. Pourtant, ce processus s’avère déterminant pour la régénération des peuplements (remplacer les individus morts), la démographie (augmenter la taille des populations), entretenir la diversité génétique (des individus issus de parents différents via la fécondation croisée à l’origine des graines) et la répartition dans l’espace. Dans le cas de notre sujet, l’endozoochorie sur des fruits charnus, une dispersion efficace va dépendre de la quantité de graines (ici des noyaux) dispersés mais aussi de sa qualité, i.e. un dépôt des graines dans les excréments à une certaine distance du parent producteur, dans des microsites favorables à la germination puis à la croissance de la plantule et au maintien de la capacité de germination des graines après leur transit intestinal. Le temps de transit constitue un facteur clé dans ce contexte à la fois quant à l’action des sucs digestifs sur les graines et surtout quant à la distance éventuellement parcourue entre la consommation et la défécation ; cette dernière dépendra aussi des rayons de circulation quotidiens de ces animaux dans la recherche de leur nourriture. Dans nos régions, les mammifères frugivores sauvages sont soit des Carnivores (voir la chronique sur ce groupe) comme renards, blaireaux, fouines, martres ou des ongulés comme des chevreuils, des sangliers ou des cerfs.
Dans les quelques études antérieures portant sur des mammifères, figure celle sur le cerisier de Sainte-Lucie conduite dans des paysages fragmentées en Espagne centrale sous climat méditerranéen : elle a démontré que les mammifères terrestres consommateurs comptent pour 2/3 des graines immigrantes dans une population de cerisier donnée, i.e. des graines venues d’autres populations plus ou moins distantes. On sait aussi que la dispersion à longue distance, un facteur de qualité important, tient souvent à un tout petit ensemble de quelques grandes espèces ; en cas de déclin ce celles-ci comme dans les habitats fragmentés, la dispersion sera fortement obérée.
Merisier

Le merisier ou cerisier sauvage s’appelle Prunus avium en latin : avium renvoie justement à l’attrait de ses fruits pour les oiseaux (avis). Il est l’ancêtre des cerisiers domestiques dont les guigniers et les bigarreautiers. Cet arbre sauvage est très répandu en plaine et jusque dans l’étage montagnard mais quasi absent en région méditerranéenne. Il recherche les sols frais et fertiles, retenant bien l’eau ; exigeant en lumière, il est capable de s’installer dans de très nombreux milieux autant en forêt que dans les accrus, ces zones de recolonisation après abandon des cultures par exemple. Sa capacité d’émettre des drageons (rejets depuis les racines) lui permet de s’étendre rapidement une fois installé à partir d’une germination. Sa floraison éclatante attire le regard car elle a lieu avant l’apparition des feuilles en avril-mai. Les fruits sont de toutes petites cerises d’un diamètre de 12 à 15mm et d’un poids de deux grammes en moyenne. Autrefois, on les surnommait amerises (contraction de cerise et amère) ce qui a donné merises. Effectivement, si elles sont riches en sucres, elles n’en sont pas moins relativement amères. D’abord rouges puis virant au noir à maturité, elles sont regroupées en fausses ombelles de 3 à 5 sur de logs pédoncules de 3,5 à 4,5cm (« queues de cerises »). Elles mûrissent à partir de mi-juin et on en trouve jusqu’à fin juillet selon l’altitude ; le plus souvent dès mi-juillet, tout a disparu. La chair juteuse adhère fortement au noyau ; au début de la maturité, ces merises restent difficiles à détacher de leurs pédoncules. Elles finissent par tomber au sol ou bien des oiseaux les font tomber en essayant de les récolter ; ainsi, même des mammifères non grimpeurs peuvent les consommer à terre.

Dans la nature, les arbres isolés en pleine campagne peuvent être le résultat d’une attention humaine qui les a conservées pour leurs fruits. Dans les fouilles archéologiques sur d’anciennes cités lacustres, on trouve régulièrement des quantités de noyaux de merises, ce qui indique que les hommes devaient les consommer régulièrement ; ils ont dû participer à la propagation de l’espèce. Souvent, on trouve des cerisiers aux fruits assez petits mais plus gros, plus sucrés et moins amers que les merises qui sont des cultivars anciens ou des hydrides ; dans l’étude allemande, les chercheurs n’ont sélectionné que des merisiers sauvages de souche pure : ils s’en sont assurés via des analyses ADN sur la majorité des spécimens retenus.

Flashés
Venons-en donc à cette étude allemande menée dans le centre du pays sur une surface de 4500 km2 dont la partie centrale est occupée par de l’agriculture intensive avec quelques taches d’éléments semi-naturels. Le merisier est présent sur l’ensemble de ce territoire et constant dans toutes sortes d’habitats, ce qui, en sus de ses fruits charnus attractifs, en fait une espèce modèle. Dans ce vaste espace, les chercheurs ont sélectionné 21 merisiers adultes avec le même volume de couronne en évitant les individus relevant en fait de cultivars (voir ci-dessus) ou ceux situés près de vergers de cerisiers cultivés susceptibles d’interférer. La sélection a été faite de manière à répartir ces 21 individus dans une gamme d’habitats allant de forêts peu gérées à des espaces agricoles de structure assez complexe avec une certaine diversité d’éléments semi-naturels (haies, arbres isolés, bosquets, …) jusqu’à des espaces d’agriculture intensive à structure paysagère hyper simplifiée. Ainsi, les merisiers étudiés se répartissent le long d’un gradient d’intensité d’usage des terres et son lot de perturbations associées.
Au pied de chaque merisier retenu, on installe un piège photo juste au-dessus du sol à 4,5m du tronc de l’arbre ; pendant 3 nuits consécutives durant la période de maturité des fruits qui commencent à tomber au sol (juin juillet en gros), ces pièges vont « flasher » les visiteurs qui seront ainsi identifiés. On relève aussi la densité de merises sur chaque arbre et celles tombées au sol. En parallèle, dans un parc zoologique, une étude est menée sur des animaux captifs en retenant les espèces observées sous les merisiers : on leur offre chacun 30 cerises et on suit leur devenir jusqu’à l’analyse des déjections (si elles ont été consommées) ; ainsi, on dispose de quelques données (en nombre limité) sur les temps de transit des noyaux de merises chez ces différentes espèces.
Enfin, autour de chaque merisier, on évalue la structure du paysage et les habitats présents à huit échelles successives emboîtées allant du rayon de 20 m autour de l’arbre jusqu’à 1 km. Cette approche va permettre de connaître précisément ce qui influence le comportement des visiteurs et l’efficacité de la dispersion selon l’échelle d’observation retenue.
Les visiteurs du soir
756 heures de suivi photographique ont permis d’identifier cinq espèces venant consommer des merises tombées au sol ; sur les 115 clichés de « flagrant délit », 41 concernaient des renards roux, 37 des chevreuils, 18 des sangliers, 12 des blaireaux et 5 des martres/fouines (indiscernables sur les clichés). Cette communauté reflète les espèces présentes sur le secteur d’étude avec le renard et le chevreuil comme espèces les plus communes ; par contre, de manière surprenante, le raton-laveur, bien installé dans cette partie de l’Allemagne, n’a pas été observé ici alors qu’il consomme beaucoup de fruits. Mais ces cinq consommateurs sont-ils pour autant des agents efficaces de dispersion des noyaux de merise ? Les résultats des tests alimentaires conduits sur des animaux captifs (voir ci-dessus) apportent des informations précieuses à deux niveaux : le temps de transit intestinal qui s’écoule entre la consommation et le rejet des noyaux et l’état des noyaux. Plus le temps de transit est long, plus potentiellement l’animal aura eu le temps de se déplacer et plus les noyaux ont des chances d’atterrir loin de l’arbre mère. Quant à l’état des noyaux, il est déterminant pour qu’ils soient en état de germer : ils doivent ressortir intacts (au moins en apparence) et s’ils sont broyés alors l’amande interne sera digérée. En général, on sait que le passage par le tube digestif (sous l’action des puissants sucs digestifs) tend à améliorer la germination en levant des états de dormance deux fois plus qu’il ne l’inhibe. Le chevreuil se distingue des quatre autres espèces par un comportement différent : sans doute à cause de son tube digestif étroit, il recrache les noyaux avant qu’ils ne s’engagent dans l’intestin et on ne les retrouve pas dans les excréments. Le tableau ci-joint résume les résultats obtenus.
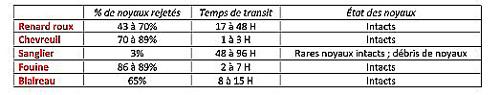
Il ressort donc de ces résultats que les cinq espèces consommatrices sont des agents potentiels de dispersion. Seul le sanglier se démarque car il apparaît plus comme un prédateur de noyaux que comme un vecteur de dispersion ; néanmoins, quelques noyaux intacts ressortent et comme le temps de transit est très long (le plus long des 5) la probabilité d’une dispersion à longue distance se trouve renforcée. Souvent, dans d’autres études, on a eu tendance à le classer à tort comme prédateur des graines des fruits charnus. Idem pour le chevreuil dont on avait omis de prendre en compte le processus de régurgitation !

3 exemples d’excréments de Carnivores avec des noyaux de merises 

Pour compléter cette évaluation, il faut maintenant considérer la taille des territoires sur lesquels circulent quotidiennement ces animaux pour rechercher leur nourriture. Celle-ci varie entre 0,05 et 1 km2 chez le chevreuil à 20 km2 chez les sangliers. Les renards et sangliers peuvent se déplacer sur près de 20 km en une nuit et parfois jusqu’à 30. Les martres et fouines circulent de 1 à 10 km par nuit et les blaireaux de 3 à 5 km. Donc en conjuguant temps de transit relativement long et rayon d’action étendu, on peut dire que ces mammifères terrestres sont d’excellents agents de dispersion des noyaux de merises.

Ajoutons que souvent ces animaux ne déposent pas leurs déjections au hasard ce qui peut augmenter les chances que les noyaux atterrissent en des microsites favorables à la germination puis au développement des plantules. Par exemple, le renard dépose souvent ses excréments sur des mottes de terre ou des taupinières (voir la chronique sur les carnivores et les autoroutes) bien en vue pour marquer son territoire ; le blaireau lui les dépose dans des trous qu’il a creusés ou pots.

De l’importance des échelles d’observation

Quatre paysages agricoles par ordre décroissant d’hétérogénéité et de proportion d’éléments semi-naturels 


Le cadre de cette étude correspond aux paysages que l’on trouve dans de nombreuses régions tempérées avec un degré d’usage des terres (et donc des perturbations) plus ou moins prononcé et des structures de paysages allant de très complexes à hyper simplifiées. Le choix délibéré dans cette étude d’explorer la structure du paysage à différentes échelles spatiales permet d’analyser ce qui impacte vraiment l’activité de ces mammifères dans le cadre de la dispersion des noyaux de merises ; et les résultats sont riches d’enseignements.

Premier résultat inattendu : les taux de visites aux merisiers sont indépendants du pourcentage de couverture forestière dans les paysages à grande échelle. Mais de études antérieures montrent effectivement que ces cinq espèces souvent réputées forestières sont capables de s’adapter à une large gamme de paysages dont des zones agricoles voire même urbaines. Par contre, le taux de visites augmente quand la proportion d’éléments semi-naturels augmente dans les paysages agricoles et baisse dans les paysages d’agriculture intensive. Ces mammifères semblent donc bien préférer malgré tout des paysages plus hétérogènes avec des mosaïques d’éléments.
A une petite échelle, quand on examine l’impact de la structure verticale de la végétation sous et aux abords immédiats des merisiers, on constate qu’elle n’influe pas sur le taux de visites ce qui confirme la prépondérance de l’influence du paysage à grande échelle. Il est donc réconfortant de voir la capacité d’adaptation de ces mammifères à des paysages très transformés par l’agriculture mais, en même temps, cela confirme l’importance extrême de conserver des paysages hétérogènes avec une part significative d’éléments semi-naturels : haies, bosquets, arbres isolés, prairies, murets de pierre, talus, … La majorité des espèces qui participent à la dispersion ayant un temps de transit long et des territoires de circulation étendus peuvent apporter un potentiel majeur d’évènements de dispersion à longue distance entre des populations d’arbres très fragmentées et isolées les unes des autres. Le cormier (voir la chronique) est à cet égard un autre exemple éloquent de ce maintien d’une circulation des graines à relativement grande échelle grâce à ces animaux. On notera aussi l’importance des Carnivores (renards, fouines, blaireaux) qui font l’objet d’une très forte pression de destruction organisée par les soi-disant « régulateurs de la faune sauvage » : il serait bien de prendre en compte cet autre aspect capital (outre la régulation des populations de petits rongeurs) pour le maintien de la biodiversité végétale.
Une seconde chronique sur la dispersion du merisier traite du rôle des oiseaux frugivores d’après une autre étude conduite par la même équipe de chercheurs.

Bibliographie
Tree visitation and seed dispersal of wild cherries by terrestrial mammals along a human land-use gradient Claudia Grünewald, Nils Breitbach, Katrin Böhning-Gaese Basic and Applied Ecology 11 (2010) 532–541
