La mort programmée des feuilles

Chaque automne, nous assistons au superbe spectacle des belles couleurs des feuillages qui précède la chute des feuilles chez les arbres (et arbustes) feuillus. Dans l’imaginaire populaire, les feuilles meurent simplement de leur belle mort à cause de la météorologie progressivement défavorable : autrement dit, cette longue agonie serait un processus subi et passif ; « les feuilles ont fait leur temps ; elles sont usées et ne serviront plus à rien en hiver ! ». Les études scientifiques nous révèlent une toute autre histoire bien plus complexe et captivante où l’arbre contrôle avec précision et moult péripéties cette longue dégradation qui conduit à la chute. Ce contrôle a une importance vitale pour sa survie à moyen terme. Tout ceci pointe une fois de plus notre méconnaissance générale de ces êtres extraordinaires que sont les arbres et de leurs capacités surprenantes.
Sénescence

L’arabette des dames, la plante annuelle modèle pour l’étude de la sénescence 
Sur cette touffe de digitale pourpre en pleine floraison, on note que les feuilles les plus basales sont en train de jaunir et sécher
Les scientifiques nomment ce processus du terme peu poétique (au regard du festival coloré qui l’accompagne !) de sénescence automnale, i.e. le vieillissement, suivi de la chute des feuilles ou abscission, autre terme très technique. En fait, le choix de ce mot n’a rien de spécifique à l’automne et aux feuilles des arbres et arbustes feuillus : la sénescence concerne une grande partie des organes de tous les végétaux qu’ils renouvellent plus ou moins régulièrement. Ainsi, même les plantes herbacées annuelles connaissent ce processus en cours de développement à propos de leurs feuilles (notamment celles du bas qui flétrissent souvent en cours de saison) mais aussi les fleurs (les pétales par exemple) et les fruits, tout comme les arbres en y ajoutant pour certains l’écorce et les brindilles ; cette sénescence peut aussi se manifester à l’occasion d’épisodes de stress comme des fortes gelées ou des sécheresses prolongées. Ceci explique le vif intérêt porté par les agronomes à ce processus qu’ils étudient à travers diverses espèces cultivées : contrôler l’abscission des fruits reste un rêve absolu pour créer de nouvelles variétés … génétiquement modifiées ! Par ailleurs, l’espèce la plus étudiée à ce propos est une petite plante annuelle sauvage, l’arabette des dames, plante modèle en génétique dont les moindres recoins du génome ont été explorés et analysés. Par contre, l’étude de la sénescence automnale des arbres n’a commencé que récemment à être explorée en détail avec un genre modèle, les peupliers, dont le peuplier tremble.

Mon précieux …

Les feuilles renferment une substance vitale pour l’arbre : la chlorophylle, le célèbre pigment qui colore les feuillages en vert ; ses molécules excitées par les photons lumineux libèrent de l’énergie récupérée et transformée en matières carbonées nutritives avec le dioxyde de carbone prélevé dans l’air via les stomates, ces minuscules orifices qui trouent la surface des feuilles. Elle procure aux végétaux verts (chlorophylliens) l’autonomie nutritive (autotrophie) avec la seule lumière et l’air, en ajoutant l’eau et les sels minéraux prélevés dans le sol via les racines. Cette précieuse molécule présente néanmoins quelques points fiables : de taille relativement grande (pour une molécule) et de structure complexe, elle est coûteuse en énergie à produire tout en étant très fragile et dure à maintenir en bon état de marche, notamment à cause des effets délétères de certains rayonnements lumineux ; elle ne peut être synthétisée que dans des cellules exposées directement à la lumière. Elle se trouve dans des organites cellulaires, les chloroplastes au sein des cellules, microscopiques grains verts qui par leur très grand nombre confèrent la teinte verte universelle des végétaux dont les arbres.

L’un des premiers signes externes les plus évidents de l’entrée en sénescence des feuilles, c’est leur virage coloré vers un jaunissement avec perte progressive de la couleur verte : ce virage coloré très connu fera l’objet d’une autre chronique spécifique. D’ailleurs, pour suivre le déroulement de la sénescence, les scientifiques prennent comme outil de mesure l’évolution de la quantité de chlorophylle dans les feuilles par rapport à son niveau estival maximal. Avec la baisse progressive de la longueur du jour, un des signaux majeurs de l’automne (voir ci-dessous), la réparation et le maintien en état de marche de la chlorophylle devient de plus en plus difficile du fait de la baisse de la quantité et de la qualité de la lumière ainsi que des apports en nutriments. Ainsi, la chlorophylle des feuilles entame un irrésistible déclin.

Feuilles de sycomore à moitié sénescentes 
Feuilles d’érable de Cappadoce dont la chlorophylle a presque entièrement disparu 
Feuilles d’érable plane tombées au sol : il ne reste que quelques minuscules îlots vaguement verts
Démolition

La démolition (catabolisme) de la chlorophylle va donc progressivement prendre le dessus sur sa synthèse et son maintien : les chloroplastes (voir ci-dessus) commencent à se dégrader et évoluent en … gérontoplastes, terme scientifique transparent pour une fois ! Mais, attention, qui dit déclin ne dit pas du tout débâcle généralisée : l’arbre a encore absolument besoin de ces feuilles tant qu’il n’a pas récupéré une grande part de leur contenu nutritif (voir la suite) et cette récupération demande de l’énergie et constitue une activité en soi. On assiste donc là à un jeu de compromis très subtil entre mort lente mais très active par ailleurs. Ainsi, dans un premier temps, seules les cellules chlorophylliennes du centre de la feuille (mésophylle) voient leurs chloroplastes se dégrader ; les cellules épidermiques dont les stomates cruciaux pour la nutrition et les cellules des vaisseaux conducteurs de sève élaborée restent intacts. Même dans les cellules dégradées, seuls les chloroplastes se décomposent et les mitochondries, ces organites clés dans la respiration cellulaire restent intactes assurant la production d’énergie à partir des nutriments récupérés.

Feuille de marronnier d’Inde attaquée par les chenilles mineuses qui creusent des galeries dans l’épaisseur des feuilles 
Une attaque généralisée induit une sénescence bien plus précoce et accélérée chez ces arbres
En fait, dans les feuilles, on trouve deux types de chlorophylles très proches, la chlorophylle a (la plus commune) et la chlorophylle b, qui absorbent des longueurs d’ondes du spectre de la lumière un peu différentes : la b, dite « chlorophylle d’ombre » est plus efficace en situation ombragée que la a dite « de plein soleil ». Dès que la sénescence s’enclenche, la chlorophylle a est convertie en b, donnant un pic de chlorophylle qui se manifeste par une teinte vert sombre qui ne dure que très peu de temps et n’est pas facile à détecter. Ensuite, d’autres enzymes entreprennent la démolition de la molécule de chlorophylle a en détachant d’abord une longue chaîne (queue phytol), puis en s’attaquant au noyau central ce qui libère l’atome de magnésium au centre de la molécule. Tous les morceaux détachés sont redécoupés et évacués de la feuille : ainsi la feuille, très progressivement va voir sa teinte verte dominante s’estomper, laissant apparaître d’autres pigments qui étaient cachés jusque là par la prééminence de la chlorophylle ou qui se sont formés (voir la chronique sur les couleurs d’automne).

Les attaques de champignons pathogènes accélèrent la chute des feuilles avant même qu’elles aient complètement perdu leur chlorophylle 
Lors de la dure sécheresse de 2019, de nombreux arbres ont vu leurs feuilles « griller » ; dans ce cas, ils ne récupèrent rien des matériaux accumulés dans celles-ci !
Recyclage

Prenons l’exemple de peuplier tremble étudié en détail (voir par ailleurs la chronique sur le feuillage « tremblant » de cet arbre et celle sur sa relation avec un grand herbivore). La démolition chimique ne concerne pas que la chlorophylle mais touche aussi les réserves nutritives accumulées pendant la saison estivale où la chlorophylle tournait à plein régime. Ainsi, en fin de sénescence, les stocks d’amidon vont être hydrolysés (démolis) en molécules de sucres élémentaires (sucrose, glucose) évacués via la sève élaborée. L’azote (N) est sans doute l’élément clé que l’arbre doit récupérer au maximum car, surtout dans les milieux aux sols pauvres ou au climat moins éclairé (comme les forêts boréales où prospère le tremble), il constitue un facteur limitant pour la croissance en tant qu’élément constitutif des protéines. Ainsi environ 90% de l’azote libéré lors de l’hydrolyse de la chlorophylle et d’autres protéines est transféré via la sève et les nervures jusque dans l’écorce où il est stocké sous forme de protéines : elles serviront au printemps suivant pour le redémarrage et le débourrement des jeunes feuillages. Ainsi, l’arbre en automne prépare t’il activement la saison de croissance de l’année suivante ! Mais le recyclage ne s’arrête pas là car une partie de ce qui est resté dans les feuilles qui finissent par tomber va rejoindre la litière au sol : là, la décomposition par la faune du sol (voir la chronique sur la litière) va libérer les éléments encore restants dont une partie pourront être récupérés par les racines des arbres quand l’eau va les entraîner en profondeur !

Feuilles mortes de tremble fraîchement tombées 
Feuilles mortes un peu plus anciennes 
Au coeur de l’hiver, l’action des décomposeurs devient nettement visible
L’arbre se trouve pris en deux feux en automne : s’il programme trop tôt la sénescence de son feuillage, il va voir sa saison de croissance raccourcie car pendant la première moitié de l’automne les feuilles poursuivent la photosynthèse et fabriquent des molécules carbonées ; s’il entame la sénescence tardivement, il risque de subir les premières gelées qui vont détruire les feuilles encore vertes sans pouvoir récupérer le précieux azote qu’elles renferment. Pris entre ces deux impératifs (carbone versus azote), sous cette forte pression sélective, l’arbre adopte donc un calendrier de la sénescence le plus adapté possible à son environnement local. Ceci pose la question du (ou des) signal déclencheur qui indique à l’arbre le moment d’entrer en sénescence.

Top départ

En fait, on constate que pour une majorité d’arbres et arbustes, le long processus de la sénescence automnale des feuillages démarre bien en amont du début d’une véritable dégradation générale du climat saisonnier. Tout comme les hirondelles partent en migration dès la mi septembre alors qu’il fait encore souvent très beau et assez chaud et qu’il y a de la nourriture disponible, les feuilles des arbres commencent extérieurement à changer de couleur sous des conditions météorologiques encore très favorables. Ceci exclut donc l’idée populaire d’un mort provoquée par le climat. La mort des feuilles est bel et bien programmée et contrôlée par l’arbre lui même, contrôle mis en place sous une forte pression sélective.

Les études sur le tremble dans le nord de l’Europe apportent des éclairages intéressants sur cette entrée en sénescence. On constate que celle-ci varie très fortement au sein de populations locales et entre arbres vivant à une même latitude mais à des altitudes différentes. Par contre, pour un tremble donné, un individu donc, la sénescence commence chaque année (sauf événement climatique exceptionnel en amont), pratiquement autour de la même date ! Un seul facteur externe perceptible peut induire une telle synchronisation : la variation de la longueur du jour par rapport à celle de la nuit, soit la photopériode. Une fois entamée, la sénescence automnale accélère sous l’effet de la baisse des températures (sans qu’il y ait de gelées). Les individus qui démarrent plus tard vont achever le processus (par la chute des feuilles) plus vite, sans doute entre autres à cause du stress imposé par la baisse des températures. La remobilisation de l’azote prend environ deux semaines (voir ci-dessus) et alors l’arbre peut se débarrasser de sa feuille vidée de son contenu !

Chez d’autres arbres comme l’érable à sucre, 90% de la variation de la date de déclenchement de la sénescence provient de variations des températures qui seraient donc le signal déclencheur ; peut être que le tremble qui vit à de hautes latitudes est une exception quant à sa dépendance envers la photopériode car elle y varie bien plus vite que plus au sud. Pour une majorité d’arbres, il semble que ce soit la combinaison photopériode/température qui est perçue comme signal déclencheur.

Erable plane en phase d’abscission 
Tapis de feuilles mortes d’érable champêtre
Calendrier
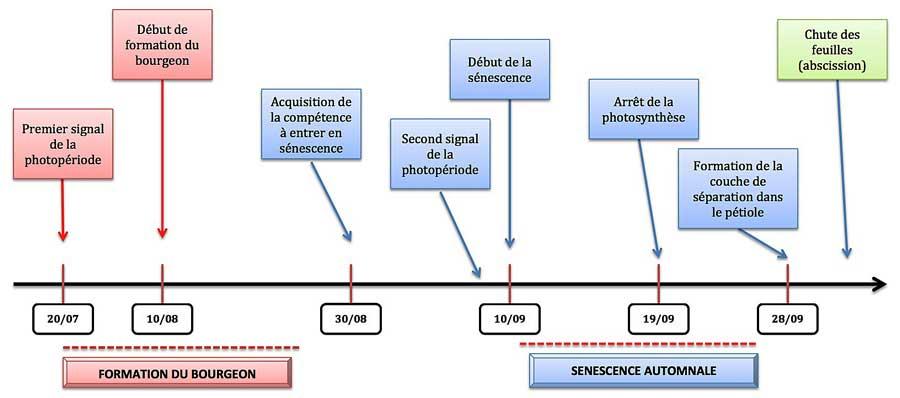
A partir des données accumulées dans plusieurs études sur le tremble, on a pu établir un calendrier type de la sénescence automnale dans le nord de la Suède et à cette occasion on a mis en évidence une relation entre le déclenchement de la sénescence et celui d’un autre processus clé dans la vie de l’arbre, la formation des bourgeons à la base des feuilles, en remplacement de ceux qui ont produit la génération printanière de feuilles. Celle-ci a lieu au cœur de l’été et passe inaperçue compte tenu de la présence du feuillage : elle aussi est déclenchée par le passage d’un seuil critique dans la photopériode qui commence à évoluer dès le début de l’été : dès l’équinoxe de fin juin passé, la longueur du jour commence à décliner doucement.

16 mai : ce rameau de tremble n’a plus de bourgeons ; ils ont donné les feuilles actuelles 
11 août : on voit des bourgeonsqui viennent de se former sur les rameaux 
12 septembre : à l’aisselle de chaque feuille il y a un bourgeon, prêt pour l’an prochain !
Il faut attendre que les nouveaux bourgeons entrent en dormance pour que l’arbre acquière une compétence à entrer en sénescence, comme une sorte de maturité acquise. Le développement des bourgeons mobilise beaucoup de nutriments dont des sucres et doit être achevé avant que l’arbre n’entame la phase de sénescence. L’importance majeure de la formation des bourgeons (sans eux, l’arbre ne peut pas produire de feuilles au printemps suivant) se reflète dans les variations observées quant aux dates : la gamme de variations est bien plus étroite pour la date de début de formation des bourgeons que pour celle de l’entrée en sénescence, très large.

Début avril : les bourgeons du marronnier s’ouvrent et produisent feuilles et fleurs 
23 avril : il ne reste plus que des écailles basales comme traces des bourgeons ! 
16 mai : sur les rameaux bien feuillés, il n’y a plus de bourgeons 
17 juillet : on aperçoit au moins un bourgeon formé mais encore réduit 
12 août : les bourgeons massifs du marronnier sont complètement formés !
Le signal de déclenchement de l’entrée en sénescence correspond à un autre seuil dans l’évolution de la photopériode qui commence à s’accélérer (début septembre). Une dizaine de jours plus tard, on observe une bascule de la source d’énergie dans les cellules : les chloroplastes cessent de fonctionner et les mitochondries prennent entièrement le relais en consommant une part des produits libérés par la démolition générale. De toutes façons, la baisse des températures (nous sommes là en Suède !) inhibe la photosynthèse ; encore dix jours plus tard, une couche de séparation se forme en travers du pétiole et la chute a lieu quelques jours plus tard.

Programmation

Un tel calendrier aussi strict, même en tenant compte des variations, suppose évidemment un fort contrôle génétique. Les études sur la plante annuelle modèle, l’arabette des dames, ont déjà identifié au moins 2000 gènes qui subissaient une activation lors de l’épisode de la sénescence ! Ces gènes sont impliqués dans un vaste ensemble de fonctions : synthèse d’enzymes pour la dégradation des macromolécules ; synthèse de transporteurs des produits issus de la démolition ; mise en place de défenses et de protection des cellules dans cette phase critique où elles commencent à décliner ; mais aussi synthèse de nouvelles protéines en lien avec les transformations cellulaires. Tout ceci confirme le statut de mort très active de la sénescence ! Sur le tremble, on a aussi mis en évidence l’activation de plusieurs dizaines de gènes associés à la sénescence dont ceux qui codent les enzymes de démolition mais aussi ceux associés aux situations de stress.

Ce processus de mort programmée s’est mis en place sous la pression de sélection évoquée précédemment ; les variations sur la date d’entrée en sénescence entre individus et entre populations apportent des possibilités d’adaptations lors de changements climatiques ou environnementaux orientés. Il rappelle un équivalent connu chez les animaux sous l’appellation d’apoptose, processus de mort programmée par lequel certaines cellules animales s’autodétruisent et se renouvellent en permanence comme par exemple les cellules de la paroi intestinale ou les cellules embryonnaires lors du développement. En dépit d’une certaine ressemblance, les deux processus semblent bien différents même si l’application de certains régulateurs de l’apoptose animale peut induire des réactions chez des plantes. Les végétaux verts ont divergé très tôt au cours de l’évolution de la lignée animale (métazoaires) et ont développé leur propre processus de mort cellulaire programmée. De plus, chez eux, elle s’applique à des organes entiers (feuilles, fruits, brindilles, ..) alors que chez les animaux elle ne concerne le plus souvent que des tissus limités !

Bibliographie
Gene Expression in Autumn Leaves. Rupali Bhalerao et al. Plant Physiol. Vol. 131, 2003
The Control of Autumn Senescence in European Aspen. Yvan Fracheboud et al. Plant Physiol. Vol. 149, 2009
Leaf senescence – not just a ‘wear and tear’ phenomenon. S. Gepstein. Genome Biology 2004, 5:212
Leaf mineral nutrient remobilization during leaf senescence and modulation by nutrient deficiency. Anne Maillard et al. Frontiers in Plant Science May 2015 | Volume 6 | Article 317
Autumn Leaf Color Development. Kim D. Coder Warnell School of Forestry & Natural Resources University of Georgia 2012
