La sève élaborée : des sources aux puits
10/01/2022 Généralement, on parle de « la » sève des plantes alors qu’en fait il y a deux très distinctes tant au niveau de leur origine, de leur composition que de leurs voies de circulation : la sève brute et la sève élaborée. Implicitement, le plus souvent, on ne fait référence qu’à la première, sans doute à cause de la visibilité plus grande, au moins chez les arbres, de ses vaisseaux conducteurs qui apparaissent sous forme de pores sur du bois en coupe ; en termes de volume circulant, elle est aussi nettement plus conséquente. La sève brute est celle qui vient des racines sous forme d’eau et de sels minéraux prélevés dans le sol et qui est ensuite distribuée dans toute la plante. La sève élaborée, elle, a une tout autre origine et circule d’une toute autre manière tout en étant aussi vitale pour les plantes. Levons donc un peu du voile qui tend à occulter cette sève élaborée dont l’appellation suggère implicitement une certaine supériorité sur la sève brute !
Photosynthèse
Si la sève brute est associée à la nutrition minérale (eau et sels minéraux) de la plante, la sève élaborée correspond quant à elle à la nutrition carbonée, i.e. aux molécules carbonées fabriquées au cours de la photosynthèse, le processus qui confère aux plantes vertes l’autotrophie, i.e. la capacité à fabriquer toutes seules leurs « aliments carbonés » et d’être ainsi autonomes (sauf la dépendance vis-à-vis du sol).

La photosynthèse prend place dans les parties des plantes renfermant des pigments chlorophylliens verts (et d’autres comme les xanthophylles ou les caroténoïdes), i.e. essentiellement les feuilles et parfois aussi les tiges si elles sont vertes. L’énergie portée par la lumière solaire (racine photo de photosynthèse) est captée par les molécules de chlorophylles concentrées dans des organites cellulaires, les chloroplastes ; là, elle est combinée avec du dioxyde de carbone prélevé dans l’air au niveau d’orifices microscopiques des feuilles (stomates) et de l’eau (apportée via la sève brute) pour produire des composés carbonés qui sont des sucres simples. Cette synthèse s’accompagne d’une libération d’oxygène.
Ces molécules de sucres seront ensuite utilisées pour la croissance, le fonctionnement (métabolisme), la reproduction et la défense contre les agressions : l’énergie emmagasinée en elles au cours de la photosynthèse est libérée via leur dégradation au cours de la respiration cellulaire, par d’autres organites cellulaires, les mitochondries : celles-ci consomment de l’oxygène et rejettent du dioxyde de carbone. Ils peuvent aussi servir à fabriquer de nouvelles molécules, autre processus qui dépense de l’énergie. Ainsi, la plante récupère l’énergie chimique stockée dans ces molécules carbonées fabriquées par photosynthèse que l’on appelle des photosynthétats ou assimilats.
Pour que la plante dans sa globalité puisse profiter de ces produits carbonés synthétisés, encore faut-il que toutes ces cellules soient ravitaillées depuis les seuls points de production que sont les feuilles (et accessoirement les tiges). C’est là qu’intervient la sève élaborée qui prend en charge ces photosynthétats et va les distribuer vers toutes les cellules. C’est en cela qu’on peut la qualifier d’élaborée : elle renferme des produits fabriqués de novo par la plante alors que la sève brute ne contient que des molécules prélevées directement dans le milieu (eau et sels minéraux) sans aucune transformation.
Sources et puits

A l’intérieur de la plante, la sève élaborée assure donc le transport des sucres depuis les organes fournisseurs capables de faire la photosynthèse ou de stocker temporairement ces sucres, les sources, vers les organes utilisateurs ou consommateurs, les puits.

La première paire de feuilles déployées prend le relais 
Les feuilles s’ajoutent et vont nourrir aussi les racines en pleine croissance
Au printemps et en été, sous nos climats tempérés, les sources majeures sont les feuilles et accessoirement la ou les tiges (et encore plus à la marge certaines parties vertes des fleurs). L’architecture et la disposition des feuilles et leur contenu en chlorophylle en font de véritables panneaux solaires dédiés à la capture de la lumière et à la fabrication de matières carbonées. Chez les jeunes plantes en cours de germination, dans un premier temps, ce sont les cotylédons, une paire de feuilles déjà formées, chargées de réserves (ainsi que des tissus nutritifs contenus dans la graine), qui se déploient et assurent la photosynthèse le temps que la première paire de vraies feuilles n’émerge et n’entre en activité. Chez les arbres ou plantes vivaces à feuillage caduque, les sucres proviennent de zones de stockage situées dans les racines ou dans la tige (le tronc pour les arbres) et remplies lors de la saison précédente ; là aussi, dès l’émergence des jeunes feuilles nouvelles et de leur mise en fonction, ce circuit de déstockage s’interrompt. Ces sites de stockage se comportent donc comme des puits majeurs en pleine saison de croissance et comme des sources tôt au printemps. Les sucres simples synthétisés dans les feuilles sont stockés le plus souvent sous forme d’amidon (une macromolécule formée de nombreuses molécules de sucres simples) : soit in situ dans les chloroplastes où a lieu la photosynthèse, soit, après transport vers des organes de stockage dans des organites cellulaires transformés, les amyloplastes (amylo = amidon).

Les puits récupèrent ces sucres pour leurs besoins métaboliques : ce sont en fait tous les tissus vivants de la plante, y compris ceux des feuilles qui elles aussi ont besoin des sucres pour fonctionner. Les plus gros consommateurs sont les organes en plein développement : les extrémités des racines qui se renouvellent sans cesse pour assurer le prélèvement d’eau dans le sol, les méristèmes i.e. les massifs de cellules qui se multiplient activement par exemple au niveau des bourgeons et assurent la croissance de la plante et la formation de nouvelles feuilles, les fleurs puis les fruits et graines au moment de la reproduction. Il faut y ajouter aussi les organismes associés avec les plantes dans le cadre d’interactions symbiotiques (comme les bactéries et mycorrhizes au niveau des racines) ou parasitaires (comme le gui par exemple sur les arbres). On notera que tous ces centres gourmands n’atteignent pas leur pic de consommation en même temps : la floraison par exemple a le plus souvent lieu après la pleine foliation ; la fructification, si elle a lieu, suit la floraison. Et puis, il y a tout le reste de la plante qui doit assurer la maintenance (dont les tissus de soutien), la défense contre les agresseurs, … et ce toute l’année pour les vivaces. A noter que les feuilles la nuit fonctionnent comme des puits car alors elles ne font que respirer et donc consomment des sucres !

Si on prend l’exemple d’un arbre, on a donc des milliers à des dizaines de milliers de sources (les feuilles) dispersées au long des branches qui doivent ravitailler autant de milliers de puits dispersés tout autant dans l’arbre depuis les racines jusqu’à la cime. Cette situation traduit bien l’extrême « décentralisation » des végétaux verts en général avec une autonomie énergétique assurée par une multitude de centres de production. Un exemple à méditer pour l’espèce humaine, championne toutes catégories de la centralisation !
En général, les sources les plus proches d’un puits donné fourniront ce site : priorité au local ! Ainsi, au printemps, les premières feuilles qui émergent fabriquent des sucres qui serviront d’abord à alimenter le développement des suivantes à proximité ; le feuillage des petites branches tend à ravitailler les branches porteuses sur lesquelles elles sont fixées ; fleurs et fruits reçoivent la nourriture fabriquée par les feuilles proches. Mais, dès que la saison bat son plein, très vite, si la plante se trouve dans des conditions favorables, la machinerie photosynthétique produit des excédents qui seront alors exportés vers les zones de stockage mentionnées ci-dessus. On estime qu’en pleine activité, chaque jour, la moitié des sucres fabriquées sont exportés pour être stockés dans les différents puits.
Passage au crible

Au-delà de la proximité immédiate des cellules productrices des sources, le mouvement des sucres fabriquées se fait via un circuit conducteur spécialisé : on parle de translocation. Elle a lieu dans un tissu conducteur spécialisé, le phloème, autrefois appelé « écorce interne » (terme trompeur !) car il se trouve juste sous l’écorce (voir la chronique sur l’écorce). D’ailleurs, le mot phloème vient du grec phloios qui signifie écorce ou enveloppe. On l’appelle aussi liber car cette partie a longtemps servi de support à l’écriture avant l’usage du papyrus ; liber est ainsi à l’origine du mot … livre !
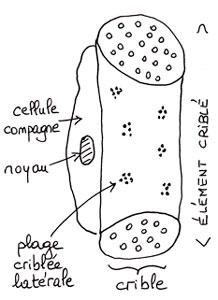
Outre des cellules banales (parenchyme) et des fibres de soutien, le phloème renferme surtout les « vaisseaux » conducteurs de la sève élaborée : les tubes criblés. Chez les plantes à fleurs (angiospermes), ces tubes sont en fait un empilement en files d’éléments cellulaires : les éléments criblés. Chacun d’eux correspond à une cellule en forme de cylindre court et trapu dont les deux extrémités aplaties portent une multitude de perforations, formant une plaque criblée ou crible ; les parois latérales portent aussi quelques petites taches de perforations au travers desquelles du cytoplasme peut passer (plasmodesme). Curieusement, ces cellules tout en ayant perdu leur noyau au cours de leur différenciation restent vivantes et gardent leur cytoplasme. Elles sont assistées par une cellule-compagne chacune, accolée contre la paroi latérale, qui possède un noyau : ces cellules compagnes ne sont pas traversées par le flux de sève mais contrôlent activement les entrées et sorties des sucres issus de la photosynthèse. D’ailleurs, cette étroite association conduit à considérer chaque couple élément criblé/cellule compagne comme un élément unité du point de vue fonctionnel (complexe conducteur). Ce système diffère radicalement de celui en œuvre dans les vaisseaux conducteurs de la sève brute et n’a pas dû évoluer historiquement en même temps mais probablement dans un second temps : l’approvisionnement en eau via la sève brute devait être la première priorité lors de la conquête du milieu terrestre par les tout premiers végétaux verts qui se résumaient peu ou prou à des axes sans feuilles.
Chez les conifères et un certain nombre d’autres groupes parmi les Gymnospermes, les tubes criblés n’existent pas : la sève élaborée circule dans des cellules criblées longues et étroites aux extrémités pointues par lesquelles elles sont aboutées. Il n’y a pas de crible aux extrémités mais des plages criblées diffuse sur les parois latérales ; les cellules associées sont différentes mais existent aussi. On pense que l’acquisition de tubes criblés propres aux plantes à fleurs a coïncidé avec le développement des fleurs.
Prélèvements in vivo

Parmi les moyens d’étude de la sève élaborée, l’un d’eux retient l’attention par son originalité : on utilise des pucerons pour prélever de la sève élaborée sur des plantes vivantes en situation naturelle ! En effet, les pucerons sont des insectes ultraspécialisés qui ne se nourrissent que de la sève élaborée des plantes riche en sucres nutritifs. Pour y accéder, ils utilisent leurs pièces buccales transformées en stylets permettant de percer et aspirer à l’instar de leurs proches cousines les punaises (voir la chronique) ; ils piquent depuis la surface et atteignent les tubes criblés du phloème qui, rappelons-le, se trouve juste sous la surface externe de la tige ; quand un tube est atteint, le puceron maintient ses stylets plantés et aspire la sève comme avec une paille : simple mais efficace ! Devant la précision redoutable de ces suceurs de sève, les botanistes ont donc eu l’idée de les exploiter comme « seringues vivantes ». On installe donc des pucerons sur la tige d’une plante étudiée ; on attend qu’ils soient en phase de prélèvement de sève : ils rejettent alors l’excédent du trop-perçu sous forme de gouttelettes de sève (le miellat). On anesthésie le puceron et on lui sectionne les stylets au ras de la tête (autrement dit, on le sacrifie !) : ceux-ci formant un tube fonctionnent alors comme un cathéter et laissent sortir la sève élaborée. Il ne reste plus qu’à prélever la sève avec une micropipette. Le gros avantage de cette méthode (outre son faible coût !) est la précision remarquable des pucerons qui ne piquent qu’un élément criblé à la fois ; on peut ainsi comparer la composition de la sève en plusieurs point d’une même tige !

Pucerons en action : le rostre est planté dans la tige et accède aux tubes criblés 

Puceron avec son rostre piqueur-suceur
Les analyses révèlent que la composition de cette sève varie considérablement selon les espèces et selon l’âge et la condition de la plante individuelle. Elle contient une part importante d’eau (80% ; voir ci-dessous à propos de la circulation) et 20% de matières carbonées. Pour l’essentiel de celles-ci, il s’agit de sucres (presque toujours du saccharose, le « sucre de table ») ; pour le reste, on y trouve des acides aminés (molécules azotées servant de briques pour fabriquer des protéines), des hormones végétales ou phytohormones, des vitamines, des éléments minéraux, des sucres combinés avec des alcools (comme le sorbitol, utilisé comme édulcorant), …

Goutte de sève élaborée en surplus rejetée par un puceron 
Feuille de bouleau couverte de gouttes de miellat
La vitesse de circulation varie elle aussi considérablement selon une foule de critères. Pour les arbres, on cite des valeurs (relatives selon les contextes) allant de 13cm/h chez des épicéas, à 48 cm/h chez les frênes ou 120 cm/h chez des ormes : autrement dit, l’ordre de grandeur tourne autour de 1m/h. Chez les plantes herbacées à croissance très rapide, la vitesse est bien plus conséquente : 2,4m/h dans des pieds de tournesols et … 6,5m/h pour des pieds de maïs !
Circulation
Dès le 17ème siècle, Marcello Malpighi, médecin et naturaliste italien, avait cherché à comprendre comment circulait cette sève dans le phloème ; il avait réalisé des expériences d’écorçage annulaire sur des troncs d’arbres : on découpe une bande circulaire d’écorce externe et du phloème juste en-dessous de manière à interrompre physiquement la circulation de la sève élaborée. Malpighi avait alors constaté qu’un bourrelet se formait en amont de la zone écorcée, signe que l’arbre continuait de grossir en épaisseur alors qu’en-dessous, la taille ne changeait pas. Cela démontrait a minima l’existence d’un flux de sève élaborée (indispensable pour construire de nouveaux tissus ligneux) descendant des hauteurs de l’arbre vers sa base et ses racines. Ultérieurement, on s’est rendu compte qu’en fait la circulation de cette sève était bien plus complexe : elle est bidirectionnelle, peut changer de direction selon les périodes et n’est pas liée à la gravité, i.e. pas seulement du haut vers le bas.

Malgré les nombreuses recherches menées sur cette sève, notamment chez les plantes cultivées, on n’a pas encore élucidé complètement son mode de circulation ; de manière inattendue, un modèle hypothétique qui date de 1927 reste le seul à peu près corroboré par diverses observations ou expérimentations. La sève élaborée circule en tout cas dans le cytoplasme des éléments criblés aboutés et passe de l’un à l’autre au travers des plaques criblées avec des ponts cytoplasmiques. Le sens de la circulation est déterminé par un gradient de pression entre les sites sources où la sève est enrichie en sucres et les puits où elle perd ces derniers. Comme les tubes criblés conservent leurs membranes vivantes, ces différences de concentrations induisent des mouvements d’eau (osmose) depuis les vaisseaux conducteurs de la sève brute (xylème) très proches des tissus du phloème. Les plaques criblées tendent à freiner et à étaler le gradient de pression qui pousse la sève en avant. Ce gradient est entretenu par le double processus de charge de sucres au niveau des sucres versus dedécharge au niveau des puits selon des mécanismes assez complexes avec des transformations des sucres simples (trioses) en saccharose notamment. Arrivée dans la tige, la sève élaborée se répartit soit vers l’amont soit vers l’aval selon la position des puits actifs du moment. La circulation montante ou descendante ne se fait pas pour autant dans les mêmes tubes criblés au sein d’un faisceau de vaisseaux. Plus un puits est actif et volumineux, plus il aura tendance à s’accaparer le flux de sève vers lui : c’est le cas par exemple des bourgeons apicaux au sommet des tiges, plus gros et entrant en premier en activité au printemps (via le processus de dominance apicale contrôlé par des hormones).
Résilience

Nous avons vu que le phloème, tissu très mince, se trouve juste sous l’écorce externe : il est donc à la merci des blessures qui peuvent affecter celle-ci : attaque d’un animal, choc externe (notamment via les activités humaines !), attaque d’un champignon parasite, … Le cambium (voir la chronique sur l’écorce), i.e. le tissu où se multiplient activement les cellules à l’origine des tissus vasculaires, pourra réparer la blessure si elle n’est pas trop importante. Par contre, des blessures majeures qui affectent notamment tout le tour de l’arbre peuvent conduire à sa mort rapide vu que les racines en recevront plus de matières carbonées (et elles n’ont pas de chlorophylle !). C’est le principe de la décortication annulaire, une méthode redoutable pour tuer un arbre sans avoir à le couper : on l’utilise notamment dans les sites protégés pour éliminer des arbres invasifs comme les robiniers ou les ailantes.

Même une petite blessure (comme lors d’une taille) peut occasionner un écoulement de la sève élaborée du fait qu’elle circule sous pression relative. Les plantes disposent de deux « pansements » pour arrêter la perte : les protéines P (P pour phloème) qui forment une masse enchevêtrée obturant la plaque criblée sous forme d’un bouchon ; le dépôt d’une substance normalement en solution dans la sève, la callose qui précipite en masse et contribue à obturer le tube crible atteint. D’ailleurs, chez la majorité des plantes, les tubes criblés ne vivent guère plus d’une saison (voire seulement quelques semaines) avant que des dépôts de callose ne les obturent complètement. Ils sont remplacés au fur et à mesure par d’autres tubes via l’activité du cambium vasculaire. Seules les « herbes ligneuses géantes » (Monocotylédones) comme les bambous, les palmiers ou les yuccas possèdent des tubes criblés restant en activité toute la vie de la plante.

A propos des écoulements de sève bien visibles en cas de blessure, il s’agit presque systématiquement d’écoulements de sève brute ! Ainsi, au printemps, quand on taille la vigne, on observe facilement des gouttes surnommées « les pleurs » mais il s’agit de la sève brute en pleine ascension du fait de la poussée racinaire. De même, le célébrissime sirop d’érable obtenu en concentrant de la sève recueillie après une incision du tronc correspond aussi à de la sève brute qui lors de son ascension tôt au printemps, avant le débourrement des feuilles, reçoit des sucres stockés dans les tissus environnants. Mais ce transport ne dure que très peu de temps. On peut l’observer facilement au début du printemps sur les bouleaux en perçant un trou vers la base du tronc et en y installant une paille ou un tube fin : on recueille ainsi la « sève de bouleau » très abondante. Ce processus permet chez ces arbres le démarrage précoce de la floraison par exemple avant la sortie des feuilles.

Bibliographie
Botanique. Biologie et physiologie végétales. S. Meyer ; C. Reeb ; R. Bosdeveix. Ed. Maloine 2004 Un excellent ouvrage de référence très didactique et richement illustré.
Applied Tree biology. A.D. Hirons ; P.A. Thomas. Ed. Wiley. 2018
An introduction to plant structure and development. Second edition. C. B. Beck. Ed. Cambridge U. Press. 2010
Botany. An introduction to plant biology. Fourth edition. J. D. Mauseth. Ed. Jones and Bartlett. 2009