Liseron des haies : un volubile qui a beaucoup à nous raconter
Convolvulus sepium

30/12/2020 Dans notre flore sauvage, le grand liseron ou liseron des haies fait figure d’ornementale améliorée tant il affiche des fleurs d’une taille et d’une blancheur remarquables sans oublier son port grimpant souvent exubérant ; les insectes pollinisateurs en profitent car il leur offre de belles ressources. On le croise de plus en plus car il a réussi à s’adapter à plusieurs environnements crées par l’homme autant en ville que dans les grandes cultures.

Volubile




Le grand liseron est une plante volubile : ses tiges s’enroulent autour des supports verticaux qu’elles trouvent. D’ailleurs, le nom latin de genre, Convolvulus, vient du verbe latin convolvere , s’enrouler (que l’on retrouve dans circonvolution). Néanmoins, tous les liserons ne sont pas volubiles mais simplement rampants au sol ou à tiges dressées comme le liseron des dunes (voir la chronique) ou le liseron cantabrique.

Liseron des dunes 
Liseron cantabrique
Contrairement à la majorité des plantes volubiles en général, ses tiges anguleuses assez fortes, souvent teintées de brun rouge, s’enroulent de la gauche vers la droite, soit dans le sens inverse des aiguilles d’une montre. La jeune pousse allongée décrit des mouvements circulaires de grande ampleur : en moyenne, en moins de deux heures, son extrémité parcourt un large cercle à la recherche d’un support : dès le contact établi, elle l’enserre et ne le quitte plus. Souvent le support est une autre tige de liseron ce qui donne naissance à ces entrelacs indémêlables typiques de l’espèce.

La plantule issue de la germination d’une graine émet à la base de ses deux feuilles initiales (cotylédons) des tiges : s’il n’y a pas de support juste à côté, elles restent couchées au sol et rampent, s’étalent tout en se ramifiant très fortement, à la recherche d’un support éventuel ; elles restent alors stériles, i.e. qu’elles ne fleurissent pas. Le mécanisme de recherche a été étudié chez les cousines des liserons, les ipomées (les volubilis horticoles), et nous y avons consacré une chronique. Les tiges qui par contre réussissent à atteindre un support peuvent fleurir.

Jeunes plants avec déjà des pousses allongées « chercheuses » 
Rapidement, le plant s’étale en ratifiant ses tiges
Ainsi, le grand liseron, selon les milieux et les hasards des rencontres, peut former aussi bien de vastes tapis étalés au sol en tous sens que des nappes escaladant la végétation basse qu’il recouvre entièrement ou bien des tresses montant à l’assaut des tiges, branches, grillages ou autres supports dressés. Toutes ces formes partagent un point commun : un grand nombre de tiges émanant de la souche souterraine (voir ci-dessous) ramifiées en tous sens et entremêlées de manière inextricable. Ces tiges herbacées pouvant atteindre jusqu’à 5m de long ne vivent pourtant qu’un an et sèchent dès l’automne, persistant un temps sur les supports dressés.



Le feuillage très fourni transforme ce fouillis végétal en une masse « étouffante ». Les feuilles grandes (jusqu’à 10cm), alternes au long des tiges, portées sur un long pétiole, présentent une forme typique en fer de flèche avec deux oreillettes basales écartées.

Plus claires en dessous, elles arborent des nervures principales palmées (comme les doigts d’une main) et se rejoignant en arceaux sur le pourtour. Leur nombre et leur taille finissent par dissimuler entièrement le réseau de tiges qui les porte. Dès l’automne, des taches foncées ou pourpres apparaissent avant le jaunissement et la mort de ces feuilles.



Forteresse souterraine
Le secret de la vitalité et du succès du grand liseron tient dans son appareil souterrain composé de gros cordons blanc jaunâtre épais (3-6mm de diamètre), charnus, très ramifiés et s’étalant parfois jusqu’à trois mètres ; ce sont des tiges souterraines gorgées de réserves : des rhizomes. Contrairement à ceux du liseron des champs, ils ne s’entortillent pas ni ne s’enroulent en spirale mais tout comme eux ils ont la capacité de s’enfoncer profondément ce qui les met à l’abri de bien des menaces y compris celle des désherbants chimiques. Tout au long de ces tiges apparaissent des bourgeons capables de donner des pousses nouvelles qui viennent renforcer le pied initial tout autour. L’autre gros avantage de ces rhizomes, c’est leur résilience à la fragmentation : le moindre fragment devient capable de redonner naissance à un nouveau pied indépendant s’il a été déplacé mais en étant une copie conforme (clonage) du pied initial. C’est ainsi que le grand liseron se retrouve plutôt favorisé par le travail mécanique du sol en surface qui fragmente ses rhizomes sans atteindre ses bases profondes ; des fragments entraînés par les machines peuvent être dispersés. Moralité : on ne se débarrasse pas aisément du grand liseron une fois qu’il est installé dans une culture : même le couvert végétal au sol ne le gène pas pour peu qu’il réussisse à s’élever sur un support comme dans les champs de maïs.


Cet appareil souterrain se met rapidement en place à partir de la plantule initiale : les premières tiges horizontales rampantes finissent par s’enfoncer dans le sol où elles s’épaississent et se transforment en rhizomes. Sur les grandes pousses se forment des rameaux courts latéraux qui s’enracinent et engendrent eux aussi des rhizomes en s’enfonçant. Ainsi, le liseron étend son emprise tel une pieuvre tentaculaire dont les bras s’enterrent pour former de nouvelles tiges qui produisent de nouvelles pousses qui … On saisit mieux ainsi comment cette espèce arrive à produire de tels massifs de végétation tant en surface qu’en volume et à être capable de recommencer année après année !

Ces rhizomes renferment des réserves nutritives accumulées via l’intense photosynthèse assurée par le feuillage opulent qui se déploie en hauteur si la plante a réussi à grimper ou à chevaucher la végétation en place. Vers la mi-juin, l’essentiel des réserves des rhizomes se trouve épuisé suite à la fabrication des nouvelles tiges et feuilles et fleurs et de la ramification des rhizomes ; tout l’été, la machine verte à l’air libre va recharger ces réserves. L’analyse chimique de ces dernières a révélé une surprise : la principale protéine de stockage ressemble très fortement aux ribonucléases, des protéines capables de découper les molécules d’ARN (on les qualifie de ce fait d’ARNases) et qui interviennent dans le processus de fabrication des protéines ; mais la protéine de réserve du liseron diffère très légèrement dans sa séquence d’acides aminés (histidine remplacée par lysine) des ribonucléases ce qui la rend inactive.

Campanette

Avec la floraison, une nouvelle vague d’abondance submerge les colonies de grand liseron : de grandes fleurs par centaines d’un blanc éclatant vont se succéder de juin à octobre. Ces fleurs ne durent pas plus d’une journée quand il fait chaud mais, une fois ouvertes, elles peuvent le rester de nuit. Elles ne s’ouvrent qu’en plein soleil ; par temps couvert ou pluvieux, elles restent repliées. D’ailleurs, seules les tiges qui ont réussi à escalader des supports et à s’exposer au maximum à la lumière fleurissent. La corolle passée flétrit très vite à la manière de celle des ipomées.
Portées chacune par un long pédoncule à l’aisselle d’une feuille, les fleurs apparaissent sous la forme d’un cône enchâssé dans deux feuilles (bractéoles) en forme de cœur, souvent teintées de brun, qui cachent le calice à 5 sépales verts plus étoits.

Bractéoles cachant en partie le calice 
Sépales vert clair du calice visibles de côté là où les bractéoles ne se recouvrent pas 
Fleurs prêtes à s’épanouir 
Profusion de fleurs en boutons qui fleuriront très bientôt
Puis, elles s’allongent et la corolle enroulée en drapeau sur elle-même se déploie en un large entonnoir évasé pouvant atteindre 8cm de diamètre avec le rebord recourbé vers l’extérieur. Cette forme caractéristique de tous les liserons est qualifiée d’infundibuliforme. Vue de dessus, on ne retrouve la trace des cinq pétales soudés que via la succession de dix plis en accordéon et d’une bande légèrement plus foncée sur le revers de chaque pétale.

Cette taille remarquable a bien entendu attiré l’attention de nos ancêtres qui lui ont accolé une série de beaux noms populaires. A cause de sa blancheur immaculée, on associé le grand liseron au culte chrétien de la Vierge Marie d’où ces noms de chemise ou manchette de Notre-Dame ; la manchette désignait un ornement de dentelle ajusté sur les poignets, évasé en forme de fleur. Mais le surnom le plus répandu, était celui de campanette (ou bien campanelle) : ce diminutif vient de campane, un ornement en forme de cloche, terme que l’on retrouve dans campanule ou campanile. Ce surnom populaire s’est aussi porté sur les narcisses avec leur fausse-corolle en tube-entonnoir ou sur des plates à bulbe proches des colchiques, les bulbocodes alpins, à fleurs évasées en entonnoir à l’éclosion. Les campanettes étaient aussi des grelots mis aux harnais des chevaux.

Fleur revolver

Etamines et anthères blanches ; noter les poils raides le long des bases élargies des filets 
Le style dépasse les anthères et le stigmate déploie ses deux lobes papilleux 
Le « barillet de revolver » 
Cage formée par les bases des filets
Il faut s’approcher et plonger le regard au fond du tube de l’entonnoir qui se rétrécit pour examiner les organes sexuels de la fleur qui atteignent à peine le sommet du tube avant son évasement. Cinq étamines blanches, aux anthères tout aussi blanches, montent, serrées autour du long style terminé par un stigmate à deux lobes. Ce dernier se déploie nettement au-dessus du sommet des anthères, une forme d’herkogamie i.e. de distanciation spatiale entre organes mâles et femelles qui diminue les possibilités d’autopollinisation. Les anthères libèrent un abondant pollen à gros grains nettement visibles à l’œil nu, caractère peu répandu chez les plantes à fleurs.

Anthère remplie de pollen 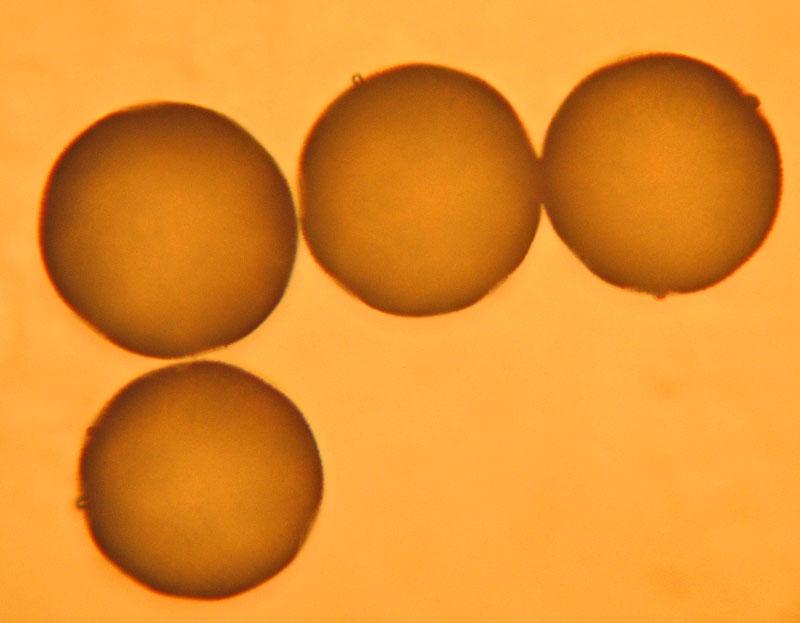
Gros grains de pollen (microscope) 
Stigmate couvert de reliefs papilleux qui captent les grains de pollen 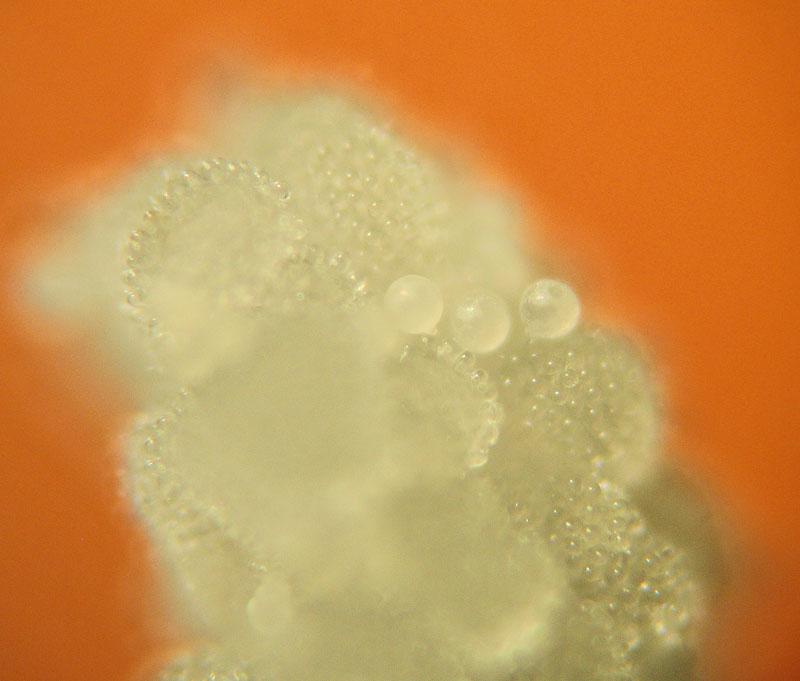
Mais le plus intéressant se trouve tout au fond du tube. Là, on voit que les filets des étamines émergent entre cinq trous formant comme le barillet vide d’un revolver. Imaginez que vous êtes un insecte butineur qui a réussi à s’insinuer au fond de ce tunnel : vous voilà bloqué et il va vous falloir être doté d’une trompe ou langue suffisamment longue et fine pour la glisser dans ces orifices et atteindre le précieux nectar.

Pour comprendre le mode d’emploi, il faut déchirer la corolle : les filets des étamines sont en fait soudés au tube de la corolle et leurs bases élargies serrées, maintenues par des poils transparents raides, ménagent entre elles comme une cage ajourée au fond de laquelle se trouve le petit ovaire ourlé par en dessous d’un gros anneau charnu orange, un nectaire sécréteur de nectar. Ainsi, les insectes pollinisateurs doivent tourner au fond de la fleur pour sonder successivement ces cinq orifices d’accès ce qui augmente le temps de visite et les chances que l’insecte porteur du pollen d’une autre fleur visitée auparavant ne le dépose sur le stigmate au cours de ces manœuvres. A noter que ces fleurs ne répandent aucun parfum : leur taille et leur couleur doivent suffire comme signaux attractifs.

Bourdons en action 
Avec la longue langue, celui-ci essaie d’atteindre le nectar 
Des méligètes noirs mangeurs de pollen accompagnent ce bourdon
Ces fleurs prodigues en pollen (très facile d’accès par rapport au nectar) et en nectar attirent de nombreux visiteurs aux premiers rangs desquels des abeilles (domestiques ou solitaires), des bourdons, des syrphes, mais aussi des coléoptères intéressés par le pollen. Il est probable que les deux bractéoles externes soient une protection pour empêcher les « tricheurs » (comme les bourdons) de percer la fleur à sa base pour accéder plus vite au nectar mais sans participer à la pollinisation. Le grand liseron fait partie des plantes ressources intéressantes pour de nombreux pollinisateurs d’autant que sa floraison généreuse s’échelonne sur une très longue période. En Europe, le liseron des haies se montre auto-incompatible, i.e. que du pollen d’une fleur qui se déposerait sur le stigmate d’une fleur de la même plante ne germe pas et ne peut pas féconder la fleur. Par contre, au Japon, où il se trouve en limite orientale de son aire de répartition très étendue, il devient autocompatible : il s’y reproduit à la fois par pollinisation croisée et par autofécondation. Ce serait une forme d’assurance de reproduction, à cause, peut-être, d’une insuffisance de pollinisateurs ou d’une compétition avec d’autres espèces de liserons locales (qui elles par contre sont autoincompatibles) qui monopoliseraient les pollinisateurs.

Les abeilles solitaires récoltent surtout le pollen 
Dispersion

Fruit vert entouré des sépales et des deux bractéoles écartées ; noter le style qui persiste au sommet 
Fruit sec avec le calice persistant
Les fleurs fécondées donnent des fruits secs, des capsules globuleuses lisses, cachées dans le calice et les bractéoles persistantes. Elles finissent par s’ouvrir progressivement et assez difficilement, libérant trois à quatre grosses graines lisses, brun foncé. Enrobées d’un tégument dur, clles-ci doivent subir les assauts du froid et de l’humidité avant de pouvoir germer dès la mi-avril suivante dans le sud de la France. Ces grosses graines ne semblent bénéficier d’aucune adaptation a priori pour un mode de dispersion ni par le vent ni par les animaux qui les récolteraient. Pour comprendre son mode de dispersion, il faut considérer les habitats naturels originels du grand liseron ; l’espèce est étroitement associée à la végétation des bords des eaux courantes ou stagnantes : forêts riveraines humides des grandes vallées alluviales (aulnaies-frênaies ; saulaies ; peupleraies à grandes herbes) ; lisières boisées humides ; roselières ; fourrés humides ; haies humides le long de fossés. On a d’ailleurs nommé Convolvulion sepium une association végétale à grandes herbes (mégaphorbiaie) des bords des eaux douces car le liseron des haies y domine en compagnie des épilobes hérissées, grandes orties, eupatoires chanvrines, consoudes officinales, … Cette relative dépendance envers l’eau libre laisse à penser que ses grosses graines doivent être transportées lors des débordements ou crues comme de nombreuses autres plantes ripicoles (des bords de rivières).




Cependant, il se pourrait que le principal mode de dispersion soit par voie végétative sous la forme de fragments de rhizomes (voir ci-dessus). Si ce dernier mode semble évident en milieu cultivé (voir ci-dessus), il s’applique aussi dans son milieu originel : les grandes crues déplacent les sédiments, érodent les berges et dégagent donc des fragments des profonds rhizomes transportés ainsi à grande distance le long des couloirs fluviaux.
Anthropophile

Roselière colonisée 
Talus-fossé en bocage humide
De son habitat riverain, le grand liseron a conservé deux exigences écologiques majeures par rapport aux sols : le besoin d’une réserve en eau utile élevée et d’une bonne charge de nutriments minéraux comme ceux déposés par les crues dans les sédiments vaseux. Il se comporte donc en espèce nitrophile ce qui l’a de facto rapproché des environnements humains perturbés enrichis en azote (eutrophiques). Ainsi, le grand liseron a conquis les régions bocagères en suivant les haies aux abords des lieux fréquentés par le bétail profitant au passage des supports crées par l’homme comme barrières, clôtures et grillages qui lui permettent d’accéder à la lumière, sa troisième exigence majeure.



Plus tard, il a commencé à entrer à la périphérie des villages, puis des villes, à la faveur des friches ou dépotoirs sur des sols argileux engorgés. On le retrouve désormais bien installé jusqu’au cœur des villes où il colonise volontiers les cimetières !


Plus récemment, profitant de la pratique grandissante de l’irrigation et des apports massifs d’engrais, il a commencé à peupler d’abord les cultures pérennes comme les vergers de fonds de vallées. Au cours des dernières décennies, sans doute favorisé de plus par le réchauffement climatique vu qu’il aime la chaleur (thermophile), il a adopté les cultures de maïs irriguées. En France et aux Pays-Bas au moins, il est devenu une « mauvaise herbe » honnie d’autant qu’il résiste très bien aux pires herbicides via ses rhizomes profonds ; il semble de plus indifférent, contrairement à nombre d’adventices des cultures, au mode de travail du sol qu’il soit griffé, disqué ou labouré !



Sur les bords de champs, ses colonies réussissent à submerger les hauts chaumes des maïs rendant impossible leur récolte. En Europe de l’Est comme en Pologne on observe la même tendance : une étude de 2016 près de Cracovie montre que 30% des champs de maïs sont concernés et qu’il recouvre près de 18% de la surface de ceux-ci ; sa présence semble favorisée par la proximité de rivières (habitat source) et des sols sableux irrigués où il peut enfoncer profondément ses rhizomes.

Vrai-Faux liseron

Le liseron des champs (Convolvulus arvensis) 
Bractéoles très peu développées laissant voir largement le calice
Jusqu’à très récemment, le grand liseron était placé dans un genre botanique différent des liserons « classiques » (Convolvulus) : Calystegia. Ce nom de genre, construit à partir des racines grecques kalix (calice) et stego (je cache), fait clairement allusion à un des caractères marquants qui avait conduit à le séparer des Convolvulus : une paire de grandes bractéoles (voir ci-dessus) recouvrant entièrement ou partiellement le calice et qui sont petites chez les Convolvulus. On peut citer encore comme autres critères distinctifs les stigmates renflés (versus filiformes), la corolle glabre (vs pubescente le long des bandes médianes des pétales) et des grains de pollen différents. Pourtant, les analyses génétiques montrent que ces « grands liserons » (26 espèces dans le Monde) forment certes un cluster mais niché au milieu de l’arbre de parentés des Convolvulus ; on les a logiquement ramené dans le genre Convolvulus comme c’est, le cas dans la nouvelle flore de référence Flora Gallica. Quant à l’épithète sepium, il signifie « des haies » (comme la vesce des haies Vicia sepium).
Par ailleurs, on a repéré depuis plus d’un siècle une forte diversité au sein de cette « espèce » qu’on a éclaté en plusieurs espèces très proches au statut incertain ; certains y verraient plutôt des sous-espèces sans compter qu’il existe de nombreux hybrides et des formes horticoles échappées de culture. Ainsi, on distingue en gros trois « espèces » : le liseron des haies (C. sepium), le liseron des bois (C. silvaticus) et le liseron douteux (C. dubius) ; le clé d’identification ci-jointe, extraite de Flora Gallica donne les critères d’identification.

La situation est rendue encore plus compliquée par l’existence au sein de l’espèce liseron des haies de formes colorées (morphes) à corolle panachée de rose pâle prêtant à confusion avec le liseron douteux à corolle elle aussi panachée de rose ! On observe apparemment des intermédiaires entre ces types panachés et ceux entièrement blancs.

Liseron douteux probable : bractéoles très recouvrantes et oreillettes des feuilles arrondies ; photographié en Vendée 
Variété rose (peut-être ?) du liseron des haies : bractéoles non recouvrantes et oreillettes aigües
Pour terminer, notons que c’est peut-être ce liseron des haies qui a suscité le nom liseron (et sa variante liset) par analogie avec des fleurs de lis blanc ! Il mérite donc bien de rester en compagnie des « vrais » liserons !

Bibliographie
FLORA GALLICA Flore de France. J-M. Tison et B. de Foucault. Ed. Biotope. 2014
Calystegia sepium – an expansive weed of maize fields near Krakow. Gala-Czekaj D, Gąsiorek M, Halecki W, Synowiec A. Acta Agrobot. 2016;69(4):1690
VARIATION OF BREEDING SYSTEM, FLORAL REWARDS, AND REPRODUCTIVE SUCCESS IN CLONAL CALYSTEGIA SPECIES (CONVOLVULACEAE). ATUSHI USHIMARU AND KIHACHIRO KIKUZAWA American Journal of Botany 86(3): 436–446. 1999.
