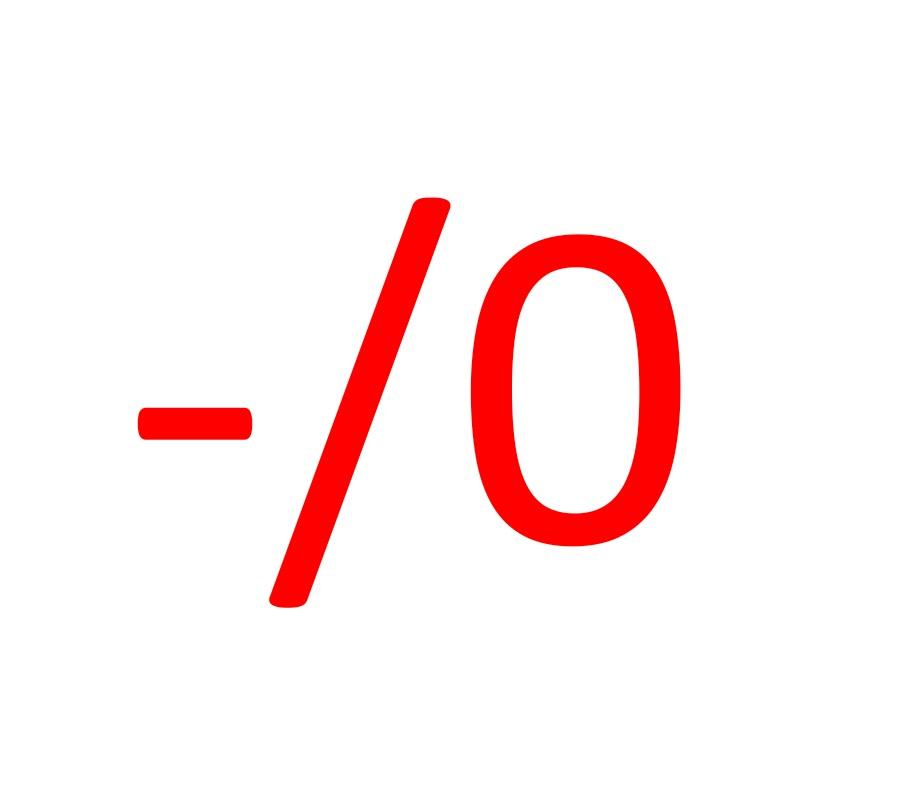Interaction fatale : faire le mort peut coûter cher !
Nous avons déjà traité des interactions entre espèces en insistant sur leur diversité : elles ne sont pas que sur le mode alimentaire (proies/prédateurs ; chaînes et réseaux alimentaires) et incluent souvent un bénéfice pour au moins un des partenaires . Dans cette chronique, nous allons aborder un exemple très surprenant d’interaction qui illustre très bien leur diversité extrême et surtout leur issue imprévisible a priori.
Amensalisme
Il existe un type d’interaction non alimentaire très particulier entre deux espèces ne correspondant à aucun des deux grands cas évoqués ci-dessus : l’une des deux espèces subit des préjudices du fait de cette interaction tandis que l’autre n’en tire aucun bénéfice ni préjudice ; c’est l’amensalisme, noté dans la codification des interactions : -/0.
L’exemple hyper classique souvent cité est celui des grands herbivores de la savane comme les éléphants qui, à l’occasion de leurs déplacements et leur recherche de nourriture, piétinent, arrachent, écrasent et même avalent toutes sortes de petites « bêtes » sans s’en rendre compte et sans en tirer a priori aucun bénéfice ; même chez nous, on a pu montrer que les troupeaux de moutons en déplacement détruisent à leur insu des nids d’oiseaux nichant au sol comme pipits et alouettes.

Une vache qui broûte avale aussi régulièrement des insectes accrochés aux herbes ! 
Un troupeau de moutons en déplacement : des dizaines de pieds qui peuvent détruire des nids au sol (alouettes, pipits, traquets, …)
Dans cette chronique, nous allons évoquer une telle interaction amensaliste mais entre deux espèces animales de même taille : un criquet et un papillon au stade chenille ; cette dernière est la perdante de l’interaction du fait de son comportement défensif très particulier.
Thanatose
Chez de nombreuses larves ou adultes d’insectes à mobilité réduite, il existe un comportement répandu d’évitement en cas d’attaque de prédateurs : faire le mort (thanatose) en se figeant sur place, en repliant ses pattes et, souvent, en se laissant tomber. Vous avez peut être déjà observé cela avec les coccinelles que l’on saisit en main. Ce comportement est censé dérouter le prédateur ou éviter qu’il ne repère sa proie si ce n’était pas encore fait.
Telle est la tactique défensive utilisée par une chenille dans cette étude (voir paragraphe suivant) : elle se nourrit sur des herbes et tend à grimper tout au bout des feuilles pour commencer à grignoter tout en revenant vers la base : ainsi, elle exploite la partie la plus tendre et la plus nutritive de la feuille. Elle se trouve de fait très exposée. Dès que cette chenille pressent une attaque de prédateur ou supposé comme tel, elle s’enroule sur elle-même et se fige, faisant donc le mort et attendant que l’alerte passe. En France, par exemple, la chenille du bombyx de la ronce est spécialiste de ce comportement : elle se roule en spirale dès qu’elle se sent menacée ce qui lui vaut le surnom d’anneau-du-diable.

Chenille de Bombyx de la ronce faisant le mort 
Guêpe de feu (chrysidé) faisant le mort ! (Cliché Y. Koide ; C.C. 4.0.)
Ainsi présenté, tout semble idéal mais le stratagème a son revers. Le plus souvent, du fait de sa position en bout de feuille, la chenille en thanatose tombe au sol et y reste un moment avant d’entreprendre la remontée sur sa plante … ce qui lui prend une bonne vingtaine de minutes à chaque fois. Autrement dit, ce comportement a un coût en termes de survie : du temps perdu à ne pas manger (et pour une chenille, il faut beaucoup et vite manger) et de l’énergie dépensée à remonter à chaque fois sur la plante. Donc, l’astuce fonctionne assez bien à condition de ne pas en « abuser » et notamment de ne pas y recourir à tort et à travers …
Prairie alpine
Ce préalable indispensable va nous permettre de comprendre l’interaction étudiée ici. Le cadre de cette étude se situe dans les montagnes du Tibet, à 3500m d’altitude, dans des prairies humides. Cadre un peu exotique mais pas si différent que cela en terme de fonctionnement et de structure de nos prairies alpines ; on pourrait donc y transposer ces résultats sans prendre trop de risque de faire de l’extrapolation abusive. Dans ces prairies hautes d’une trentaine de centimètres en moyenne, dominent deux grands groupes d’espèces : des laîches, plantes proches des graminées (ici genre Kobresia) et des potentilles qui représentent à elles deux 90% de la biomasse de tiges et de feuilles de ce milieu. Deux insectes herbivores (consommateurs de feuilles) dominent très nettement dans ces prairies : un criquet (Chorthippus fallax).
et la chenille d’un papillon nocturne (Gynaephora alpherakii ). A partir de la mi-août et jusqu’à fin septembre, ces deux espèces dominent largement avec des densités de 60 individus par m2 pour le criquet et de 20 à 100 chenilles/ m2. D’ailleurs, ces chenilles étant toxiques, elles posent des problèmes aux éleveurs de yaks, moutons ou bovins qui exploitent ces alpages !

En cage
Les chercheurs ont mis sur pied un protocole original pour contourner la difficulté quasi insurmontable de ne s’appuyer que sur des observations directes. Ils ont installé dans ces prairies des cages de taille réduite posées sur une surface de végétation. Quatre types de cages sont mis en place : des cages avec que des criquets (10/cage), ou que des chenilles (10), ou avec les deux réunis (10 et 10), ou aucun des deux (cages vides d’herbivores). On recrée ainsi des microcosmes presque naturels où la densité des protagonistes se rapproche de celle observée (50 individus au m2).
On suit sur presque deux mois l’évolution des uns des autres (poids, taille, transformations pour les chenilles, évolution de la biomasse végétale sous cage) tout en recueillant des informations directes sur les comportements adoptés par les chenilles lors des contacts avec les criquets. Et c’est là que se situe justement le problème : les criquets très actifs sautent beaucoup au milieu des herbes ce qui déclenche chez les chenilles le réflexe de thanatose : elles réagissent comme si les criquets étaient des prédateurs, ce qui n’est absolument pas le cas. Néanmoins, il existe d’autres espèces « ressemblantes » d’orthoptères comme certaines sauterelles qui, elles, peuvent consommer des insectes en plus des végétaux.
Ainsi, dans les cages où chenilles et criquets sont réunis, les chenilles tombent en moyenne 3,5 fois par heure aux périodes où elles sont actives alors que, lorsqu’elles sont seules, les chutes « tombent » ( !) à 1,25 par heure ; en effet, les mouvements des herbes par le vent peuvent aussi susciter cette réaction.
Prix fort
Autrement dit, c’est le manque de discrimination dans la réponse défensive des chenilles qui pose problème : une sorte d’excès de zèle qui va leur coûter cher comme en attestent les résultats.
Les chenilles en compagnie des criquets passent en moyenne 1060 minutes par jour à se nourrir, contre 1920 minutes quand elles sont seules : elles perdent donc 45% de leur temps à remonter ! Par contre, les temps de nourrissage des criquets restent identiques dans les deux situations. Les chenilles qui côtoient les criquets se nymphosent (tissage d’un cocon et transformation en chrysalide) en moyenne 4 jours plus tard : leur cycle de vie est donc plus long.
En termes de survie, le nombre de criquets restés en vie dans les cages n’est pas affecté par la présence de chenilles, bien qu’ils se nourrissent comme elles, essentiellement des feuilles de laîches ; la biomasse végétale est telle qu’il n’y a pas de compétition même avec ces densités. Par contre, le nombre de chenilles survivantes et transformées en cocons au bout des deux mois des expériences est significativement plus bas dans les cages mixtes.
Donc, clairement, les chenilles subissent une pression négative forte à cause de la seule présence des criquets qui les « dérangent » et suscitent le comportement de thanatose ; elles se nourrissent moins, s’épuisent à remonter sur leurs plantes après une chute : elles grandissent donc moins vite et leur survie est affectée de manière significative.
L’intensité de cet effet collatéral inattendu a de quoi surprendre mais il tient à plusieurs « circonstances aggravantes » : ces deux espèces recherchent les mêmes plantes nourricières et sont donc amenées à se côtoyer ; la densité des criquets est élevée ce qui crée des mouvements permanents dans l’herbe ; et enfin, les deux espèces présentent les deux mêmes pics d’activité journaliers : en milieu de matinée et en milieu d’après-midi. En fait, les chenilles, dans cet environnement, ne sont qu’assez peu confrontées à de réels prédateurs (des moineaux surtout) et sur-réagissent aux criquets inoffensifs !
Effet cascade
Les conséquences de cette interaction ne s’arrêtent pas aux seules chenilles et à leur survie : la végétation subit elle aussi un impact. La biomasse totale de végétaux sous chaque cage a été estimée à deux reprises pendant l’expérimentation : sous les cages mixtes (chenilles + criquets) elle était 122 à 84% plus élevée que sous les cages avec chenilles seules. Donc, les perturbations subies par les chenilles réduisent la consommation de végétaux (elles sont particulièrement gloutonnes !) recherchés. Les laîches se développent alors beaucoup plus au détriment des autres plantes comme les potentilles.
Par contre, la biomasse sous cages mixtes (chenilles + criquets) est plus faible que sous les cages avec criquets seuls : les criquets ne réussissent pas quand même à complètement empêcher l’impact herbivore des chenilles !
Clichés
L’interaction de type amensaliste entre criquets et chenilles change donc subtilement le fonctionnement de cet écosystème en modifiant les rapports entre espèces végétales dominantes ; les chenilles seules auraient un impact considérable car leur capacité de consommation et d’exploiter la ressource végétale est élevée. D’autres interactions du même type entre espèces de taille proche sont progressivement mises à jour mais pas faciles à débusquer et il est encore moins facile d’évaluer leur impact quantitatif. On s’éloigne donc encore plus du « tout alimentaire » comme seul mode de relation entre espèces différentes !
Ces exemples montrent que les fameuses notions galvaudées « d’équilibre entre espèces » ou de «nature harmonieuse » en prennent un coup : pas étonnant car il ne s’agit ni plus ni moins de considérations purement anthropomorphiques teintées de créationnisme religieux !
Bibliographie
Grasshoppers amensalistically suppress caterpillar performance and enhance plant biomass in an alpine meadow ; Xinqiang Xi, John N. Griffin and Shucun Sun. Oikos 122: 1049–1057, 2013