Les mousses : deux plantes en une (épisode 1)

15/11/2021 De tous les végétaux terrestres, les mousses, et leurs cousines les hépatiques, sont les plantes les moins bien connues et surtout les plus mal comprises. Les idées reçues ne manquent pas à leur égard et ont été longtemps entretenues par la vision hiérarchisée et fixiste du monde vivant et de l’évolution : les mousses sont toujours présentées comme des végétaux inférieurs, primitifs, … des sous-plantes en quelque sorte qui n’auraient servi qu’à permettre l’émergence des plantes supposées supérieures. Leur petite taille a beaucoup contribué à cette vision complètement fausse et anthropomorphique. Un autre obstacle majeur s’oppose à leur appréhension pour ce qu’elles sont : la compréhension de leur cycle de vie radicalement différent de celui de toutes les autres plantes terrestres. Les profanes sont souvent incapables de décrire comment est faite une mousse au niveau de ses feuilles, de ses tiges, … Par contre, beaucoup retiennent d’elles ces sortes de longues tiges fines surmontées d’une capsule qui émergent des tapis ou des coussinets : des sporogones. Mais qui sait que les dits-sporogones sont en fait des plantes à part entière littéralement perchées sur la plante verte feuillée. Ces deux plantes superposées correspondent à deux stades très distincts du cycle de vie des mousses très surprenant dans son déroulement. Ce premier épisode vous incite à entrer dans les arcanes de ce cycle de vie et de découvrir plus en détail ces fameux sporogones perchés sur les mousses : qui sont-ils, à quoi servent-ils ?

Deux générations

Les mousses s’inscrivent dans une grande lignée dite des plantes terrestres (même si certaines d’entre elles sont retournées secondairement en milieu aquatique), laquelle a émergé de la lignée verte (les êtres chlorophylliens) qui s’était développée dans les océans avec entre autres les algues vertes. Les scientifiques nomment cette lignée qui a conquis le milieu terrestre depuis la mer les embryophytes, les plantes à embryon. Dans le cadre de leur reproduction sexuée, la fécondation ou fusion de deux cellules sexuelles donne une cellule-œuf qui se développe en un embryon multicellulaire qui va être « conservé » et abrité un temps par la plante.
Quatre grandes lignées ont divergé suite au début de cette conquête du milieu terrestre :
- les bryophytes au sens large : les anthocérotes, les hépatiques et les mousses (les bryophytes au sens strict)
- les lycopodes et alliées (voir la chronique sur cette lignée tout aussi méconnue)
- les fougères et les prêles
- les plantes à graines (spermatophytes) qui incluent les plantes à fleurs (Angiospermes), les cycas, les Ginkgos, les Conifères et les Gnétales avec l’énigmatique Welwitschia (voir la chronique).
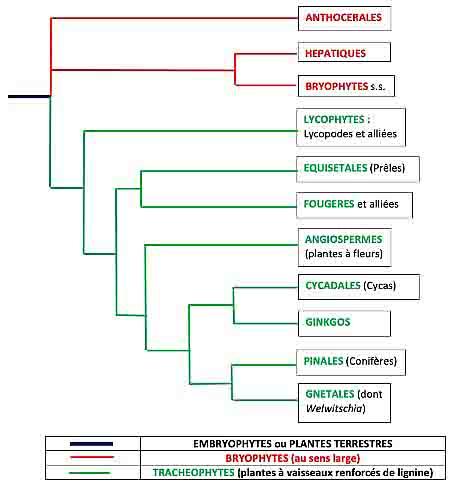
Les trois premières lignées se caractérisent par un cycle de vie où alternent deux générations représentées chacune par un « être », une « plante » différente. L’une des deux fabrique et libère des spores, éléments unicellulaires dont le noyau ne contient que la moitié des chromosomes (cellules haploïdes ; de haplos, simple) ; on appelle cette plante le sporophyte, le « porte-spores » (sporo = spore ; phyto = plante) ; d’ailleurs, on qualifie souvent l’ensemble de ces trois ignées de ce terme informel de plantes à spores. Ces spores germent et donnent naissance à une nouvelle « plante » sur laquelle vont se développer deux types d’organes sexuels producteurs de cellules sexuelles ou gamètes : on donne donc le nom de gamétophyte (le porte-gamètes) à cet organisme. Il porte soit des anthéridies, organes sexuels mâles, qui libèrent des spermatozoïdes nageurs (appelé anthérozoïdes) ou des archégones, organes femelles, qui renferment des cellules femelles immobiles, les oosphères. Les anthérozoïdes libérés nagent dans la fine pellicule d’eau qui entoure les organes sexuels et vont féconder les oosphères, donnant naissance à une cellule-œuf, à l’origine d’un embryon qui, en se développant redonnera une nouvelle plante à spores, un sporophyte. Et le cycle est bouclé !

Chez les fougères et les lycopodes (voir la chronique), on retrouve ces deux « plantes » qui alternent sous deux formes très différentes : la plante feuillée (le lycopode ou la fougère tels que nous les voyons) représente la « plante à spores ». Elle porte des sporanges, petits organes ronds dans lesquels se forment des spores qui sont libérées et dispersées. Ces spores arrivent au sol et germent pour donner des micro-plantes parfois très réduites, des prothalles porteurs des organes sexuels (anthéridies et archégones) ; ces prothalles indépendants (le plus souvent) sont donc les gamétophytes, les plantes à gamètes. On a là un cycle où alternent deux générations mais l’une des deux, celle des sporophytes, les plantes feuillées, s’y trouve surreprésentée.

Chez les plantes à graines et encore plus chez les plantes à fleurs, la génération des gamétophytes se trouve hyper-méga-réduite à quelques cellules incluses si bien que 99,99% de leur cycle de vie est représenté par le sporophyte qui est la plante feuillée. Autrement dit, dans ces trois lignées (lycopodes, fougères et plantes à graines) se manifeste une tendance à une évolution vers un cycle où le sporophyte, la plante feuillée, occupe une place prédominante.
A contre-sens

Les mousses et les hépatiques quant à elles ont évolué dans une direction diamétralement opposée : elles sont les seules plantes terrestres actuelles avec un cycle de vie doublement différent : d’une part, la plante feuillée est le porte-gamètes (gamétophyte) via ses organes sexuels portés au bout des tiges ; d’autre part, suite à la fécondation des cellules femelles restées dans leurs organes femelles (archégones), les embryons se développent en un porte-spores (sporophyte) qui va pousser directement sur sa « mère » sous la forme d’un sporogone : une longue soie dressée terminée par une capsule dans laquelle se formeront les spores. Après leur libération et leur dispersion, ces spores germent sur un substrat et vont donner des pieds feuillés porte-gamètes. Ainsi, le cycle des mousses (et des hépatiques) se rapproche plus d’un cycle équilibré entre deux générations avec une dominance malgré tout de la plante porte-gamètes, soit l’inverse de ce qui se passe chez toutes les autres plantes terrestres (voir ci-dessus). Ceci souligne la profonde divergence qui sépare les mousses et hépatiques des autres plantes terrestres, divergence qui ressort par ailleurs dans de nombreux autres domaines dont les stratégies de gestion de l’eau radicalement différentes. De ce fait, elles représentent une option évolutive tellement différente que les taxer d’infériorité relève du ridicule total : dans leur voie, elles ont tout autant réussi que les plantes à fleurs souvent citées comme le graal de la réussite évolutive.

Chez les mousses, la germination des spores donne d’abord naissance à une plantule qui se développe en un réseau filamenteux très diffus, le protonéma, ramifié ou non sur lequel vont ensuite se développer des tiges feuillées porteuses des organes sexuels.
Notons que la description ci-dessus se réfère à la majorité des espèces mais il existe des variations notables avec la réduction plus ou moins marquée de l’une des deux générations. Ainsi, les espèces adaptées à des milieux très secs tendent à réduire le porte-spores qui perd sa tige et se résume à une capsule. D’autres au contraire, réduisent fortement les porte-gamètes et on connaît même des cas extrêmes où la reproduction sexuée se fait directement sur la plantule (protonéma) ! Preuve supplémentaire, s’il en était besoin, que le potentiel évolutif des mousses reste très actif !
Le fait que ce cycle si particulier se retrouve aussi bien chez les mousses que les hépatiques et les anthocérotes a longtemps conduit à penser qu’elles dérivaient toutes d’un ancêtre commun unique ; or, les données moléculaires ne valident pas cette hypothèse : autrement dit, ce type de cycle aurait évolué au moins deux fois indépendamment.
Pied et soie
Nous allons donc maintenant consacrer le reste de cette chronique à l’exploration du porte-spores, le sporogone, l’élément des mousses qui attire le plus le regard. L’épisode 2 s’intéressera aux porte-gamètes, les mousses feuillées. L’architecture du sporogone, en dépit des innombrables variations, reste globalement simple : une longue soie dressée, jamais ramifiée, sans aucune feuille, ancrée au sommet d’une tige ; au sommet, une capsule de forme très variée qui est l’équivalent du sporange des fougères (mais ici, il est unique) et dans lequel se fabriquent les spores.

Le pied du sporogone, rétréci ou bulbeux s’enfonce au milieu d’un groupe de feuilles au sommet de la tige feuillée du porte-gamètes femelles : il correspond à l’embryon qui s’est développé dans un organe sexuel femelle (archégone). Il est entouré d’un tissu composé de cellules de transfert appartenant au porte-gamètes. Sa structure rappelle celle des suçoirs des plantes parasites ou haustorium (voir l’exemple des cuscutes) ; pour autant, on ne sait pas clairement si ce tissu est essentiel pour la nutrition du sporogone ou s’il ne sert qu’à son maintien mécanique. En tout cas, le sporogone dépend exclusivement du porte-gamètes pour assurer son ravitaillement en eau et nutriments puisqu’il n’est pratiquement pas chlorophyllien. Juste après la fécondation, l’archégone, l’organe sexuel femelle forme un sac qui enveloppe l’embryon puis le jeune sporogone. Sous l’effet de la pression engendrée par la croissance de ce dernier, ce sac de protection se déchire en deux : la moitié inférieure reste au pied tandis que la moitié supérieure va devenir une sorte de chapeau ou coiffe (calyptra) qui va recouvrir la future capsule.

Sporogones avec l’embryon basal développé 
Ici, on a dégagé l’embryon caché par des feuilles 
Parfois, la base prend une forme bulbeuse
La croissance du sporogone se fait selon un mode surprenant. Au tout début de sa croissance, le jeune embryon progresse via une cellule apicale qui se divise mais s’arrête très vite de fonctionner pour laisser place à l’entrée en service d’un massif de cellules (un méristème) qui va se diviser à la fois vers le bas et vers le haut (croissance intercalaire) ; les cellules au-dessus s’organisent en une capsule et celles en-dessous donnent la soie.

Coiffe à angle droit sur une funaire 
De face, on dirait un bec ! 
Le rostre au sommet de l’opercule soulève la coiffe qui se fend
La croissance de la soie élève la capsule tout en laissant l’embryon originel entouré des feuilles. Cette « tige » est pleine et n’est jamais ramifiée ; parfois, on y trouve un faisceau central de longues cellules minces conductrices d’eau (voir l’épisode 2). Parfois, les cellules de « l’écorce » de cette soie avec des parois irrégulièrement épaissies induisent une torsion ou une courbure de la soie qui facilite la dispersion des spores, notamment chez les petites espèces vivant sur le sol nu (terricoles).

Coiffe en place 
.. et tirée 
.. pour révéler l’opercule surmonté d’un rostre
A noter que dans les lignées basales des mousses dont les célèbres sphaignes, ces mousses des tourbières, il n’y a pas de soie faute de croissance intercalaire ; c’est le porte-gamète qui élabore un faux-pied (pseudopode) qui soulève ainsi la capsule quand même !
Capsule

La soie se termine donc par un organe renflé, la capsule dans laquelle vont avoir lieu les divisions à l’origine des spores. Curieusement, cette capsule porte des stomates (absents des soies), ces micro-orifices qui régulent l’entrée d’air (et du dioxyde de carbone nourricier) et la sortie de la vapeur d’eau en s’ouvrant ou en se fermant chez les autres plantes terrestres. Leur ressemblance superficielle ne signifie pas pour autant qu’ils sont équivalents. En effet, sur les capsules, très rapidement, ils restent ouverts en permanence ; on pense qu’en fait qu’ils n’assurent qu’une fonction mécanique en participant, à maturité, à la déshydratation des parois de la capsule qui va permettre son ouverture. Dans les lignées basales des mousses, ces stomates sont absents ce qui suggère qu’ils ne seraient apparus qu’après la divergence des mousses et de manière indépendante de ceux des fougères ou des plantes à graines.
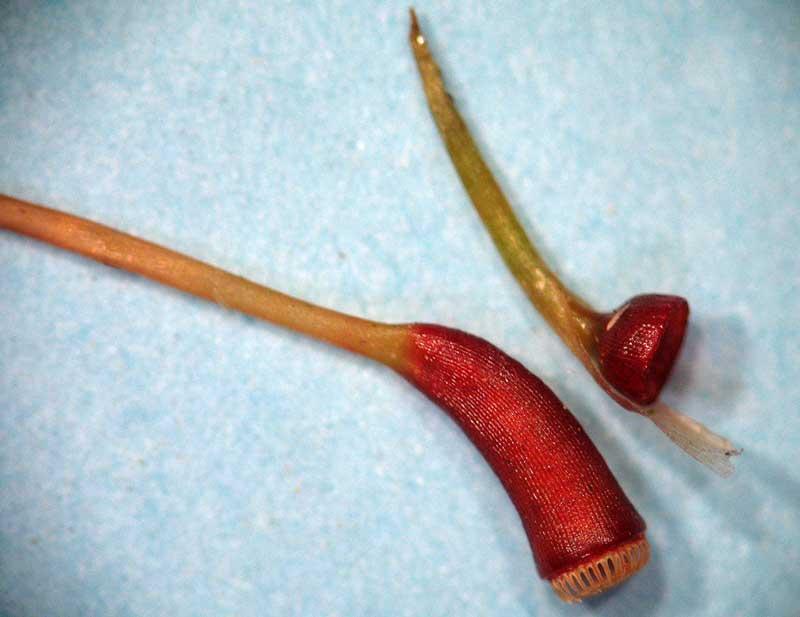
Capsule avec péristome et opercule (avec coiffe) détaché 
Péristome formé d’une rangée de dents 
Péristome à dents qui s’écartent vers l’extérieur 
Epiphragme perforé qui ferme la bouche de la capsule 
Dents du péristome
Le sommet de la capsule est couronné par un opercule, un couvercle, surmonté éventuellement d’une coiffe (voir ci-dessus). La chute de l’opercule ouvre la bouche de la capsule mûre ce qui laisse la possibilité aux spores d’être libérées notamment quand la capsule est secouée par le vent. Dans de nombreuses lignées de mousses, on assiste au développement d’une structure faite d’un ou deux cercles concentriques de dents (péristome) qui régulent par leurs mouvements la libération des spores. Comme toujours chez les mousses, ce sont les forces différentielles engendrées par les alternances hydratation/déshydratation sur les parois cellulaires d’épaisseurs différentes ; elles peuvent ainsi se resserrer vers l’intérieur ou au contraire vers l’extérieur selon les genres. Encore mieux chez certaines mousses, une membrane perforée à sa périphérie (épiphragme) vient fermer la bouche de la capsule assurant ainsi une libération graduelle des spores comme avec une salière. Mais chez d’autres mousses, c’est la capsule toute entière qui se fend en long selon des lignes de manière irrégulière.
Dispersion
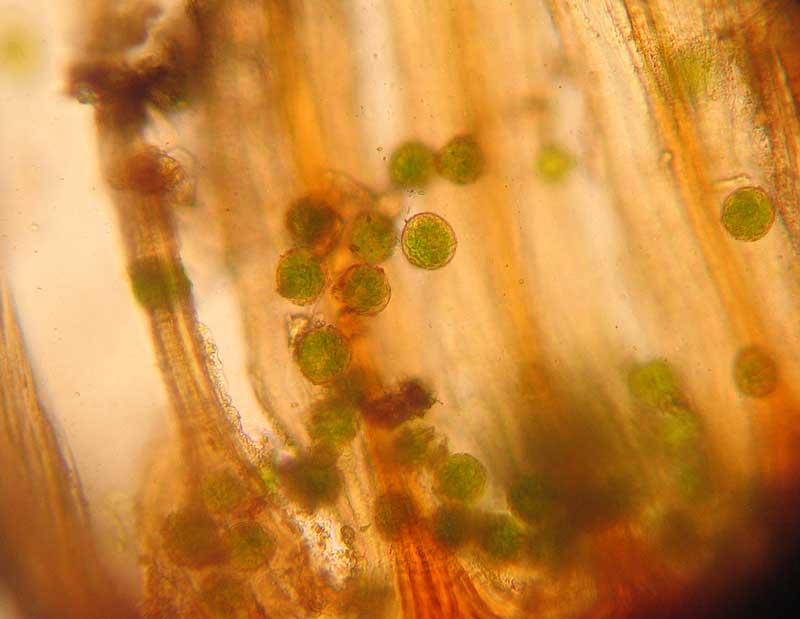
Spores au milieu des dents du péristome 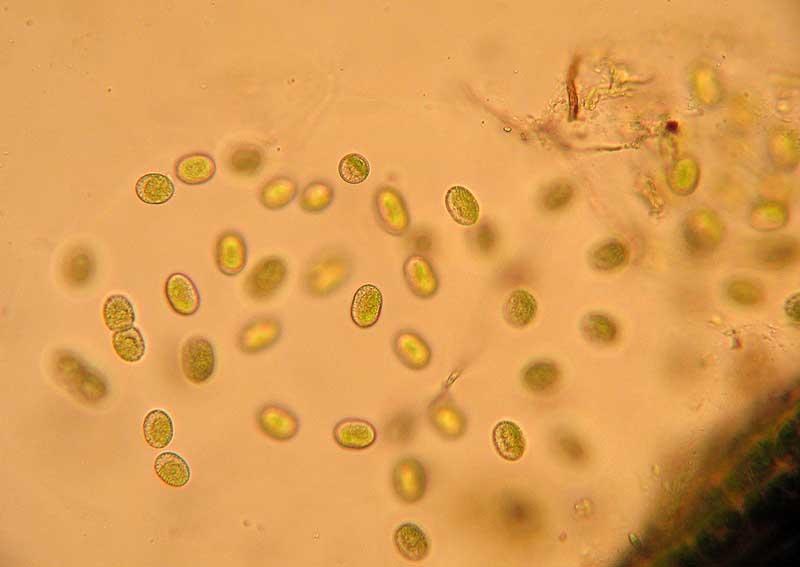
Spores : éléments unicellulaires qui n’ont rien à voir avec les graines des plantes à fleurs
On aura saisi d’après la diversité des structures présentées ci-dessus qu’il existe une forte pression de sélection sur la manière de libérer les spores ; effectivement, de cette étape va dépendre leur capacité ensuite à se disperser dans l’environnement et permettre la conquête de nouveaux milieux ou d’échapper à des conditions locales devenues hostiles. Les spores microscopiques sont néanmoins très résistantes car leur membrane (ce sont des cellules) est imprégnée d’une substance qui résiste tant aux stress mécaniques que physiologiques, la sporopollénine.

Soie courbée qui facilite la dispersion vers le sol (espèce terricole) 
Soie très courte chez les espèces de milieux très secs ou terricoles
Le plus souvent la dispersion est passive par le moindre courant d’air apte à transporter à grande distance ces spores unicellulaires ultralégères. L’élévation de la capsule au sommet d’une soie prend ici tout son sens en augmentant les chances de prise en charge des spores par le vent. Son raccourcissement chez les espèces terricoles (sol nu) associé souvent à la torsion de la soie correspond à une autre stratégie : coloniser les espaces adjacents dénudés plutôt que de partir loin et d’avoir peu de chances de trouver de tels sites. Chez d’autres mousses, des modifications des parois peuvent induire un resserrement de l’ouverture et limiter la sortie massive des spores : libérer d’un coup toutes ses spores comporte un risque du tout ou rien selon les conditions météorologiques de l’instant ; les disperser à petites doses sur plusieurs jours ou semaines augmente les chances de croiser des conditions favorables à la dispersion à longue distance. Comme les autres plantes à spores dont les fougères, ceci explique pourquoi nombre d’espèces de mousses ont souvent des répartitions très vastes quasi planétaires : l’extrême légèreté des spores permet des transports à très grande distance sur des centaines ou des milliers de kilomètres sans doute.
De leur côté, les sphaignes, lignée basale vraiment divergente à plus d’un titre, ont développé un système sophistiqué d’implosion de la capsule avec éjection de l’opercule et projection des spores après accumulation interne de gaz ! Ceci rappelle la dispersion explosive des graines de certains fruits des plantes à fleurs (mais les spores ne sont pas du tout des graines !).

Capsule verte 
Capsule sèche : des renforcements en long accentuent les tensions lors du dessèchement
Mais il y a encore mieux : une famille de mousses, les Splachnacées, a développé un dispositif de dispersion via des insectes à la manière de la pollinisation entomophile des plantes à fleurs. Ces mousses hautement spécialisées ne vivent que sur des excréments ou des cadavres d’animaux, autrement dit des milieux très éparpillés dans la nature. Leurs capsules renflées ont des couleurs vives attractives (blanches, jaune vif, rouges) et leurs soies émettent des composés volatiles qui imitent l’odeur du substrat en cours de décomposition. Elles attirent ainsi des mouches qui vont véhiculer les spores en se rendant vers d’autres sites favorables !

Jeunes capsules en début de formation 

Vous n’avez pas décroché en route et vous voulez connaître aussi « l’autre plante » qui compose une mousse : la plante feuillée ou porte-gamètes, le gamétophyte des botanistes ? Alors, rendez-vous dans l’épisode 2 pour la découvrir.

Bibliographie
INTRODUCTION TO BRYOPHYTES. A. Vanderpoorten ; B. Goffinet. Cambridge UP 2010
