Les souches : des pépinières pour jeunes arbres

14 12 2020 Dans la chronique « Les souches : du bois mort plein de vie », nous avons exploré la biodiversité animale très riche, la faune dite saproxylique, associée aux souches en décomposition en forêt. Mais la contribution des souches au maintien de la biodiversité ne s’arrête pas au seul monde animal : ce bois mort en décomposition sert aussi de substrat de développement pour de nombreux végétaux verts dont les mousses et hépatiques qui les habillent souvent d’un beau manteau vert ! Dans les forêts montagnardes, les forestiers ont depuis longtemps repéré que les souches, tout comme le reste du bois mort pourrissant au sol (bois coupé non exporté, troncs tombés, branches, …) servent de lits de germination et de développement des plantules puis des jeunes conifères (épicéas et sapins le plus souvent) : elles participeraient donc au processus de renouvellement des individus dans les peuplements ou régénération naturelle. Quelle est l’importance des souches vis-à-vis de cette régénération des conifères et qu’est ce qui les rend aptes à assurer ce rôle surprenant ?

Résineux

Dès 1904, le botaniste allemand A. Engler (1844-1930) avait observé dans les forêts presque naturelles de Bohême les bienfaits du bois mort en décomposition au sol pour la régénération des épicéas. Ainsi dans ces forêts, il n’est pas rare de trouver des alignements de jeunes arbres ayant tous germé sur un tronc pourri au sol devenu invisible après sa décomposition complète. A partir des années 1930, cette interaction a été largement documentée dans les forêts boréales de Scandinavie puis dans de nombreux écosystèmes forestiers d’altitude de l’Hémisphère Nord. Dans les forêts pluviales de la côte ouest des USA, on a démontré que la régénération des essences dominantes, l’épicéa de Sitka et la pruche hétérophylle, se confine presque entièrement sur du bois mort en décomposition au sol ; de même au Japon dans des forêts très arrosées d’Hokkaido avec un sous-bois touffu de bambous nains, on observe la même dépendance de la part des résineux dominants locaux.


En France, ce processus concerne surtout l’épicéa commun et le sapin blanc le plus souvent mais aussi le mélèze d’Europe et dans une moindre mesure certains pins montagnards. Leur point commun est de partager des petites graines ce qui implique des plantules très grêles au départ et de ce fait très fragiles et donc sensibles à la compétition de la part de la végétation du sous-bois. Un des facteurs clés semble être l’accès à la lumière pour les plantules, besoin qui s’accentue avec l’altitude croissante

La végétation luxuriante des forêts subalpines constitue un frein à la régénération 
Jeune Sapin de Douglas dans une forêt auvergnate : dans son pays d’origine, il utilise beaucoup ce genre de microsite
Cette capacité à coloniser ainsi le bois mort au sol leur procure un avantage compétitif au sein des forêts humides avec une végétation de sous-bois très développée ; celui-ci s’exprime d’autant qu’on monte vers l’étage subalpin avec l’accumulation de neige en hiver sur les pentes qui détruit les plantules trop faibles au sol.

Pessières alpines 

Microsites

L’épicéa commun est l’essence la plus étudiée de ce point de vue : il produit des graines ailées très légères libérées par l’écartement des écailles des cônes qui pendent aux branches, donnant une véritable pluie de graines entraînées plus ou moins loin, au hasard, par le vent ou par simple gravité. La production varie énormément d’une année à l’autre avec des années « fastes » où, massivement, les arbres d’un site donné produisent de grandes quantités de graines. Et pourtant, au dessus de 1500m dans les Alpes, sa régénération naturelle devient souvent difficile qualifiée de capricieuse par les forestiers. Dans les années 1990, on estimait ainsi que plus de 15000 hectares de pessières (forêts d’épicéas) en Savoie étaient touchés par ce déficit de régénération induisant une désagrégation du manteau forestier. Les graines peuvent certes germer sur les plages de sol dénudé mais ces semis disparaissent rapidement à cause de la neige et de l’humus brut épais (matière organique mal décomposée).

Microsites rocheux : ici des mélèzes et des épicéas 
Moins favorables pour la croissance ultérieure !
En fait, pour avoir des chances significatives d’obtenir des plantules par germination puis ensuite des jeunes plants qui grandissent et deviendront capables de prendre le relais des individus morts, les graines doivent atterrir en des endroits spéciaux réunissant un ensemble de conditions favorables : des microsites. Ceux-ci doivent être capables d’intercepter et de retenir les graines emportées par le vent, de leur fournir un substrat humide et nutritif permettant leur germination et doivent se trouver en position surélevée pour échapper à l’ombrage de la végétation du sous-bois et accéder à la lumière et à la chaleur et moins subir l’écrasement de la neige.

Ces microsites peuvent être le pied de rochers émergeants, les troncs couchés au sol, les bosses induites par les chutes d’arbres (chablis) qui soulèvent la terre avec leur couronne racinaire déterrée et les souches. Dans les forêts exploitées, cas de figure prédominant, les souches sont souvent de très loin les microsites les plus nombreux, les arbres morts sur pied étant souvent évacués avant de se casser et de tomber au sol ainsi que les chablis.



Quelques chiffres illustrent l’ampleur de cette importance des souches : dans les Alpes italiennes, 43% des souches portent des plantules ; en Pologne, alors que les gros bois morts au sol dont les souches ne couvrent que 4% de la surface, près de la moitié des plantules poussent dessus avec une densité de plantules 20 fois supérieure à celle sur le sol nu. Dans les Alpes du nord, 41% des 309 souches inventoriées portaient des plantules mais avec de fortes variations selon la végétation du sous-bois ; dans 4/5 des sites étudiés, les souches étaient de loin les sites les plus utilisés pour la régénération.
Avantages de nature
Cette forte prévalence des souches comme microsites de régénération leur a valu d’être qualifiées de « germinatoires » par un écologiste suisse ! Comment expliquer cette propriété remarquable ?
Les souches conservent leurs racines et se trouvent près du sol : de ce fait, elles retiennent plus facilement l’humidité capitale pour les plantules d’arbres ; leur surface de coupe ou de cassure à l’horizontale intercepte plus facilement la pluie ou l’eau de fonte de la neige qui pénètre d’autant mieux que la décomposition avance. Si le cœur de l’arbre était déjà attaqué par une forme de pourriture, la dépression centrale deviendra rapidement une cuvette propice à l’infiltration de l’eau.

Jeune sapin installé sur une racine morte de la souche 
Le dépôt de litière créé un microsol sur la surface de coupe et maintient l’humidité
La position légèrement surélevée place les plantules en meilleure place pour capter la lumière par rapport à la strate herbacée ; elles ne subissent pas non plus la compétition directe des systèmes racinaires des autres plantes dans le sol. De même, la neige s’y accumule moins et y reste moins longtemps. Par contre, plus les souches sont hautes (notamment celles, naturelles, issues de cassures de troncs), moins elles seront favorables : plus exposées au vent qui soufflera plus facilement les graines déposées ; tendance à s’écrouler quand la décomposition est avancée ; partie supérieure plus sèche ; … Assez rapidement pour les souches ombragées, mousses et hépatiques les colonisent souvent en « moquettes » vertes : certes elles retiennent l’humidité, mais elles finissent par devenir un obstacle à l’installation.
La surface horizontale, d’autant plus rugueuse qu’elle provient d’une cassure, favorise l’interception des graines sans oublier la présence de résine ; ceci vaut notamment dans les pentes fortes où les surfaces de coupe des souches conservent leur horizontalité ! Elles retiennent aussi plus de litière de feuilles mortes ce qui facilite la formation d’un humus suspendu.
La coupe du tronc expose des tissus plus tendres et nutritifs : l’écorce interne et l’aubier (voir l’autre chronique sur les souches) qui se décomposent en premier et plus vite ; les graines arrivées sur ces zones germeront mieux et enfonceront plus vite leurs racines dans ces couches concentriques verticales.

Le moindre creux devient idéal 
Les racines de ce jeune épicéa ont suivi verticalement le bois de printemps plus tendre des cernes
Enfin, il y a les champignons mangeurs de bois, super-décomposeurs (voir la chronique Les fossoyeurs du bois mort), qui s’installent rapidement et transforment le bois: outre les changements nutritifs considérables positifs qu’ils induisent (voir ci-dessous), ils semblent tisser des associations avec les jeunes racines des plantules en formant des mycorhizes qui, en retour, améliorent considérablement la survie et la croissance des jeunes plats. Les substances chimiques qu’ils libèrent éloignent par ailleurs un certain nombre d’agents pathogènes.

Diversité de souches

Souches de coupe en plein soleil taillées relativement haut 
Souche basse en sous-bois 
Souche creuse : la cavité devait exister à l’origine 
Souche haute un peu basculée (chablis coupé) ; noter les polypores très présents
Les souches peuvent être créées par l’homme lors des coupes d’éclaircie (dans les peuplements en cours de développement et notamment ceux issus de plantations), les coupes partielles dans la gestion en futaie jardinée répandue en montagne ou des coupes à blanc quand elles concernent l’ensemble d’une parcelle. Ces souches se reconnaissent facilement à leur surface de coupe bien franche et droite même si souvent apparaît au centre une zone déchiquetée là où l’arbre a cassé en phase finale d’abattage. Il s’agit le plus souvent d’arbres en bonne santé exploités pour leur bois ou présentant au plus quelques débuts d’attaques périphériques par des galeries d’insectes. La coupe se fait au plus bas ce qui donne des souches très basses, proches du sol. Les racines maintiennent un temps la souche en vie ce qui lui conserve son humidité originelle ; rapidement, la souche meurt avant de commencer à se décomposer (voir ci-dessous).

Les souches naturelles, elles, résultent de la cassure d’arbres morts sur pied ou déjà très endommagés par des attaques de parasites ; d’une hauteur moyenne de 0,5m à 2m, elles offrent une section déchiquetée, très irrégulière et du coup plus propice à l’interception des graines et à la décomposition ; mais leur hauteur limite la colonisation (voir ci-dessus). Elles diffèrent profondément des précédentes par leur histoire avant la cassure : les arbres morts sur pied peuvent rester longtemps en place debout perdant progressivement leurs grosses branches et se transformant en « chandelles » verticales (snags des anglo-saxons). Ainsi, 30 ans après un épisode de sécheresse conjugué avec une attaque de scolytes sur un peuplement d’épicéas, 40% des arbres morts étaient encore debout ! Les champignons ne manquent pas de commencer à les attaquer ce qui les fragilise mais il faudra souvent un événement brutal comme un fort coup de vent pour les casser. Certains individus peuvent ainsi se casser au bout de 4 à 8 ans selon aussi leur diamètre. En tout cas, pendant tout ce temps, le bois mort a commencé à se dessécher en position verticale : la souche issue de la cassure sera radicalement différente et ne se décomposera pas du tout à la même vitesse. Pour que de telles souches deviennent des lits de germination efficaces, il faudrait que le tronc casse peu après la mort de l’arbre sur pied. Parmi les souches de coupe évoquées plus haut, certaines concernent de tels arbres morts sur pied que les forestiers tendent à évacuer suite à ces épisodes massifs de mortalité de plus en plus fréquents avec la crise climatique.

De jeunes feuillus peuvent aussi s’installer comme ce bouleau au milieu des plantules de sapins 
Souche de coupe investie par de jeunes sapins
Tout ceci explique que des souches proches de même âge (même année de coupe ou de tempête par exemple) peuvent avoir des évolutions très différentes sans oublier l’importance de leur situation par rapport à la lumière (voir l’autre chronique sur les souches).
Décomposition

Un autre facteur va diversifier les souches : leur rythme de décomposition. Les germinations et la survie des plantules en pratique ne se fait guère que sur des souches ayant atteint un stade avancé de décomposition. Les changements alors en cours favorisent de plus en plus l’installation des jeunes plants : apparition de craquelures, de fissures qui créent une surface rugueuse retenant mieux les graines ; diminution de la densité du bois facilitant la pénétration des racines ; ouverture des vaisseaux du bois qui deviennent autant de micro-tuyaux permettant l’infiltration de l’eau ; colonisation par les mousses qui entretiennent l’humidité et facilitent les interactions entre les champignons décomposeurs et les jeunes racines (mycorhizes).

L’écorce se détache assez rapidement suite aux attaques des insectes saproxyliques 
Les souches hautes en fin de cycle risquent de s’écrouler et de ruiner la survie des colonisateurs
Cette phase optimale se situe en moyenne 30 à 60 ans après la mort de l’arbre ; cependant des germinations peuvent avoir lieu bien plus tôt dans les 10 à 20 ans suivant la mort si l’arbre avait déjà des zones dégradées en son cœur. En phase finale, quand la souche se couvre totalement de mousses et voit s’installer des fougères ou des plantes à fleurs, la compétition rend le milieu moins favorable.

Stade très avancé : le coeur se fragmente 
Souche en fin de vie, engloutie par la litière 
La mousse estompe les silhouettes ! 
L’installation de plantes herbacées (séneçons) et d’arbrisseaux freine le développement des jeunes arbres
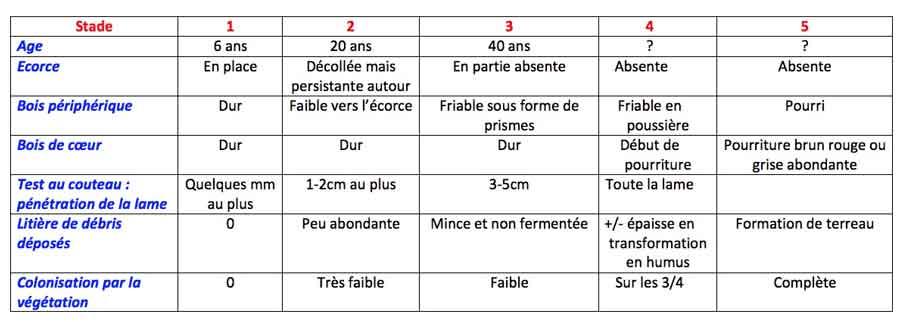
Concrètement, sur le terrain, les écologistes forestiers distinguent cinq phases de décomposition permettant de donner un âge très approximatif à une souche d’épicéa, présentées dans le tableau ci-dessous. On estime en effet que la vitesse de décomposition du bois peu varier d’un facteur 20 selon les conditions ! Pour faire ce diagnostic, à part un bon sens de l’observation, il faut juste vous munir d’un couteau pour tester la dureté relative du bois au niveau de la coupe ou de cassure.
Puits nutritif
L’aspect extérieur de la souche et ses qualités physiques évoluent certes mais les changements peut être les plus importants mais invisibles concernent sa composition chimique sous l’action des champignons décomposeurs et de l’intense activité microbienne : les nutriments disponibles sont un élément clé pour la survie et la croissance des plantules. Des études scandinaves ont minutieusement suivi le devenir des éléments chimiques au cours du processus de décomposition sur une période de 40 ans après la coupe. Sur cette période, les souches perdent presque 80 % d e leur carbone, du fait de la dégradation de la matière organique : ce carbone est passé en grande partie dans l’atmosphère sous forme de CO2 mais il a été libéré très progressivement, bien moins vite que si on brûlait par exemple les souches après les avoir déracinées ! Par contre, on assiste en parallèle à un phénomène inverse très surprenant : une nette augmentation de la quantité d’azote, un élément clé pour la nutrition des jeunes arbres ainsi que du phosphore, autre nutriment clé. Autrement dit, de manière contre-intuitive, les souches deviennent de plus en plus nutritives : elles se bonifient avec le temps alors qu’elles sont en pleine décomposition ! Mais d’où viennent ces éléments en plus ? L’apport par les précipitations est très faible dans les régions étudiées et on ne peut pas invoquer le sol qui se trouve « à côté » ni les racines qui sont mortes. Les responsables seraient les champignons décomposeurs qui investissent les souches : ils y développent des longueurs considérables de filaments (mycélium) souvent organisés en éléments ressemblant à des racines (rhizomorphes ou cordons mycéliens). Via ce réseau considérable en volume et surface, ils envahissent non seulement la souche mais aussi ses environs et ils vont donc prélever au niveau du sol environnant des composés azotés ou phosphorés qu’ils accumulent dans la souche, leur bastion central. On parle de translocation d’éléments chimiques. La proximité spatiale du sol favorise ces transferts. Les souches fonctionnent donc comme des réservoirs à éléments chimiques nutritifs à long terme avec l’aide des champignons.
Un autre élément nutritif retient l’attention : le potassium ; on le retrouve dans la formule chimique de nombreux engrais NPK : N pour l’azote (Nitre autrefois) , P pour le Phosphore et K pour le Potassium (Kali autrefois). Dans le sol, quand cet élément est libéré suite à la décomposition de la matière organique de la litière, il est très vite entraîné en profondeur ou en surface par les écoulements d’eau à cause de sa forte solubilité dans l’eau : on parle de lessivage. Or, sous les climats montagnards arrosés, ce lessivage intense appauvrit vite les sols en ce précieux élément. Et bien, dans les souches, la quantité de potassium se maintient mieux là aussi via la translocation par les champignons qui limitent ainsi les fuites.
Tous ces éléments accumulés ne seront libérés très lentement qu’au delà de cette période de 40 ans, redistribuant ainsi ces ressources mises en réserve et permettant une bonne croissance des jeunes plants, sans oublier l’important facteur de l’humidité retenue par le bois pourri.
Match de pourris !

D’avant en arrière : chablis déraciné, tronc au sol et volis pas encore séparé (une future souche haute !)
Pour parachever cette apologie des souches comme « germinatoires » à résineux, nous allons les comparer avec un autre élément important du bois mort au sol : les troncs ou morceaux de troncs ou grosses branches tombées au sol. Diverses études ont suivi la régénération des épicéas sur les souches en parallèle de celle sur les morceaux de troncs immédiatement à côté issus le plus souvent de la cassure de l’arbre mort ayant engendré la souche.

Troncs au sol 

En Suède (forêts boréales), on observe une plus forte proportion d’arbres matures poussant sur des souches que sur les troncs morts bien que les très jeunes plants soient plus nombreux sur ces derniers car la décomposition y démarre plus tôt (contact étendu avec le sol) ; les souches se rattrapent donc en termes de meilleures conditions de survie et de croissance. Dans une forêt résineuse semi naturelle au Japon, on trouve quatre fois plus de plantules de conifères sur les souches par rapport aux troncs morts. En Europe centrale, dans de vieilles pessières d’altitude, les densités des plantules sur les microsites sont de 0,2/m2 sur des sites sans bois mort (tas de terre ou base de rochers), 5,5/m2 pour les troncs morts au sol et … 19,5/m2 pour les souches ! Souches : vainqueur par k.o. technique ! Cette disproportion s’accroît avec l’altitude avec des densités six fois supérieures dans les pessières subalpines. La composition et notamment la hauteur de la végétation au sol influent beaucoup sur cet avantage ; ainsi par exemple, l’abondance de fougères sur les troncs morts au sol limite encore plus l’installation des plantules. La part des souches augmente aussi fortement dans les forêts exploitées d’où on tend à exporter les troncs morts tombés au sol, pratique en cours d’abandon mais qui a prévalu longtemps en Europe centrale ; localement, il y a aussi les exploitations « sauvages » illégales de bois au sol qui accentuent encore plus de déséquilibre en faveur des souches.

Tant que le tronc de ce chablis sera « suspendu » au-dessus du sol, il se décomposera peu 
Jeune épicéa sur un tronc pourri au sol
Quel spectacle !

Je ne peux pas terminer cette chronique sur les souches sans évoquer ma fascination pour ces dernières justement depuis que j’ai commencé à observer ce phénomène de nurserie perchée : d’abord en Auvergne dans les forêts de la Chaîne des Puys ou du Sancy et encore plus dans les pessières subalpines des Alpes. Ces souches colonisées soit par des plantules, soit par de jeunes arbres en croissance, offrent des spectacles hautement symboliques : le passage d’une génération à l’autre et la renaissance de la vie à partir de la mort ! On ne va pas faire comme certains auteurs à succès de l’anthropomorphisme et y trouver des intentionnalités de la part des arbres mais plutôt y voir de belles illustrations de l’action des ingénieurs de l’environnement (voir la chronique sur cette notion) que sont notamment les champignons, élément-clé de toute cette histoire !

Une fois l’attention attirée sur ce processus, on acquiert un œil plus aiguisé quand on se promène en forêt : prenez le temps dès maintenant de vous arrêter dès que vous voyez une souche pourrie ; n’hésitez pas à quitter le sentier pour l’inspecter de près ; touchez la, humez la pour mieux cerner l’état de la décomposition ; évitez de la démolir d’un coup de pied ce qui revient à détruire brutalement un microcosme en pleine maturation plein de vie.

Epicéas échassiers installées sur d’anciennes souches disparues depuis 

Alors, avec ce nouveau regard, vous saurez mieux repérer certaines scènes hautement photogéniques. Il y a ainsi les arbres à échasses sur des fantômes de souches : ces arbres ont poussé au départ sur une souche, donc en hauteur et ont développé des racines latérales plus superficielles qui sont passées par dessus la souche ; une fois celle-ci complètement décomposée, il se crée parfois un vide qui donne à la base de l’arbre une allure singulière ! Que dire de ces jeunes arbres perchés sur des souches hautes et qui surplombent le sol de la forêt de leur jeune insolence ? Et encore plus de quelques rares scènes ahurissantes : ce jeune épicéa vaillant qui pousse tout en haut d’un vieil adulte au tronc étêté ou bien, spectacle rarissime, ce tronc couché suspendu au dessus du vide et colonisé par une armée bien alignée de jeunes épicéas.

Jeune épicéa installé au sommet d’un adulte décapité 
Il a l’air en forme même au coeur de l’été 2020 en pleine canicule !
Pour moi, de ces scènes émane autant d’exotisme que bien des paysages ou lieux vantés comme « à couper le souffle » et situés de préférence à l’autre bout de la planète !

NB. Ce sujet des souches a fait l’objet d’une émission enregistrée par Christophe Noiseux, animateur du magazine H2O sur France Bleu Pays d’Auvergne ; vous pouvez écouter ce reportage effectué dans la chaîne des Puys en cliquant sur ce lien.
Bibliographie
Density and height structure of seedlings in subalpine spruce forests of Central Europe: logs vs. stumps as a favourable substrate. Bace, R., Svoboda, M. & Janda, P. 2011. Silva Fennica 45(5): 1065–1078.
Phosphorus and base cation accumulation and release patterns in decomposing Scots pine, Norway spruce and silver birch stumps. M. Palviainen et al. Forest Ecology and Management 260 (2010) 1478–1489
Régénération en altitude de l’épicéa Picea abies sur les souches dans les Alpes françaises. P Gensac. Ann. Sci. For (1990) 47, 173-182
Spruce regeneration on woody microsites in a subalpine forest in the western Carpathians. Bujoczek L., Bujoczek M., Banaś J., Zięba (2015). S. Silva Fennica vol. 49 no. 3 article 1337. 21 p.
When does dead wood turn into a substrate for spruce replacement? Zielonka, Tomasz Journal of Vegetation Science 17: 739-746, 2006
Tree regeneration on rotten wood and on soil in old-growth stand. J. Szewczyk & J. Szwagrzyk Vegetatio 122:37-46, 1996.
