Les taupins, grands maîtres du click-and-jump
Elateridae

20/01/2022 Le statut d’animaux à six pattes et à quatre ailes articulées sur un exosquelette confère aux insectes des capacités locomotrices qui nous déconcertent, nous qui raisonnons avec nos archétypes de mammifères bipèdes ou quadrupèdes. Parmi ces capacités, celles qui touchent au saut ne cessent de nous époustoufler dès lors qu’on les rapporte à leur taille : le bond d’un criquet ou d’une sauterelle (voir la chronique sur les orthoptères), les sauts de puce imprévisibles des cercopes (voir la chronique), … ne cessent de nous étonner. Mais tout cela n’est pas grand-chose face aux étranges sauts des taupins, des coléoptères (voir la chronique sur ce groupe) très répandus sous nos cieux. Quand ces insectes se retrouvent sur le dos, ils réussissent, par une prouesse physique dont on n’a pas encore entièrement déchiffré les rouages, sans les pattes et sans élan, à sauter en l’air en une fraction de seconde et à retomber face ventrale contre terre. Le tout s’accompagne d’un petit bruit sec, un clic typique, signe d’un mécanisme de type ressort ; ceci leur vaut une foule de noms populaires évocateurs comme marteau, maréchal, forgeron, tape-tape ou tape-maillet ainsi que le nom anglais très imagé de click-beetle ! Après une rapide présentation de ces coléoptères, nous nous concentrerons donc sur ces fameux sauts.

Elatéridés

Les adultes sont floricoles 
Les taupins forment une famille de plus de 9000 espèces dans le monde, les élatéridés, remarquablement homogène du point de vue morphologique ce qui permet de distinguer assez facilement un taupin d’un coléoptère d’une autre famille. Par contre, au niveau des espèces, la tâche devient très difficile justement du fait de cette homogénéité d’aspect : plus de 200 espèces sont répertoriées en France ! Voici donc le portrait-robot général des taupins adultes (voir la chronique sur les coléoptères pour les termes techniques propres à ce groupe*) :

Noter la profonde charnière (en arrière du pronotum) qui saute aux yeux 
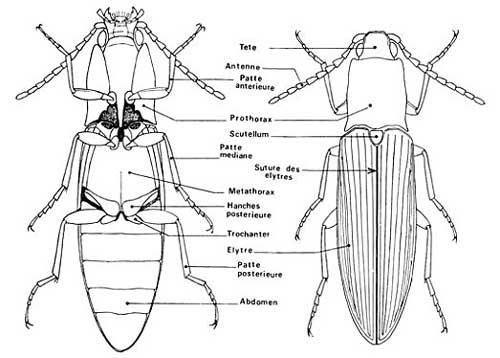
Extrait de la référence 1
- Corps de forme allongée, étroite, légèrement aplati, avec les bords parallèles
- Élytres* coriaces et résistants, striés, nettement courbés vers leur extrémité terminale
- Pronotum* arrondi en dôme avec le bord avant très arrondi sous lequel la tête est encastrée ; dans chaque angle postérieur, il y a souvent une épine orientée vers l’arrière ; par-dessous, on trouve une sorte de plaque en forme de « menton » qui cache la base des pièces buccales
- Pronotum séparé des élytres par un sillon profond qui marque une charnière d’articulation majeure
- Antennes filiformes ou pectinées à 11 articles souvent logées complètement au repos dans un profond sillon sous le pronotum
- Sous le thorax, une longue pointe dirigée vers l’arrière se loge dans une cavité du segment suivant (organe du saut : voir ci-dessous)
- Pattes courtes et peu développées ramenées sous le corps au repos
- Coloration terne : bruns, noirs, gris ; quelques espèces avec les élytres rouge vif
- Taille allant de 0,5 à 2cm (en France).

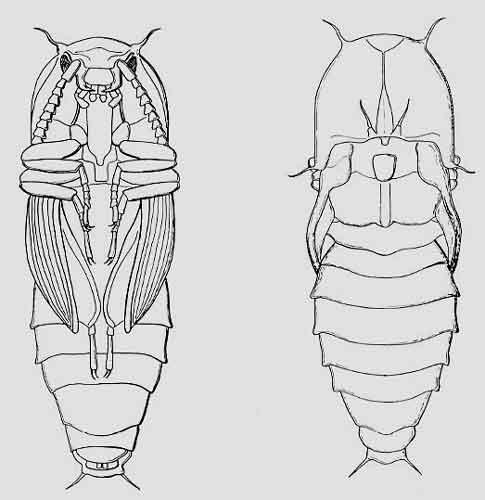
Taupin rongeur : dessus 
Charnière entre le pronotum en bouclier et les élytres 
Dessous 
Elytre écarté 
Aile postérieure membraneuse pliée sous l’élite 
Gros plan sur la pointe engoncée dans sa cavité entre les pattes
Les adultes volent et fréquentent souvent les fleurs où ils se nourrissent du pollen ou du nectar. Les larves très allongées ont trois paires de pattes minuscules ; de couleur brune à jaune, on les surnomme souvent vers jaunes ou « vers fil de fer » (à cause de leur peau très durcie). Elles se nourrissent soit de racines de plantes dont des plantes cultivées (comme les pommes de terre ou les céréales), soit pour la majorité de bois mort en décomposition (troncs creux avec du terreau notamment dans les têtards : voir la chronique) ou d’écoulements de sève (saproxylophages). Le nom de taupin vient du mode de vie des larves : au 15ème siècle, les taupins étaient des fantassins employés pour creuser des tranchées ! Les larves se transforment en pupes* souvent au bout de plusieurs années. Celles-ci ressemblent fortement aux adultes notamment au niveau du pronotum typique.
Saut de carpe

Venons-en donc maintenant au cœur du sujet : le saut si particulier des taupins adultes. Quand l’un d’eux tombe sur le dos (ou qu’un observateur amusé en place un ainsi dans le creux de sa main pour le plaisir de le voir faire), il a du mal à se remettre dans le bon sens à cause de la forme de son corps et de ses courtes pattes. Le plus souvent, il n’essaie même pas et, tout au contraire, il replie ses pattes (et aussi ses antennes quand il existe des sillons où elles peuvent se loger : voir ci-dessus). Il entreprend une cambrure de son corps au niveau du profond sillon qui sépare le pronotum des élytres, lesquels recouvrent les deux autres segments du thorax rattachés à l’abdomen (voir la chronique sur les coléoptères). Le pronotum (avec la tête engoncée) et le reste du corps couvert par les élytres font alors un angle pointé vers le haut et son corps ne s’appuie plus que sur le dos du pronotum bombé et sur le milieu aussi bombé des élytres. Il maintient cette posture immobile, tendue, pendant 1 à 3 secondes. Puis brusquement, il se détend et inverse la cambrure de son corps en une fraction de seconde (de 14 à 18 millisecondes !) avec un bruit sec typique, un clic nettement audible, comme quelqu’un qui claque des doigts. Cette détente qui fait sursauter l’observateur non initié projette l’insecte en l’air à la verticale où il peut tourner sur lui-même avant de retomber presque au point de départ, sur son ventre et ses pattes … ou pas ! S’il ne réussit pas et se retrouve de nouveau sur le dos, il recommence plusieurs fois jusqu’à retomber du bon côté et peut alors s’enfuir (peut-être) avec ses pattes. Des mesures effectuées sur des taupins acajou, une espèce commune brun luisant, donnent une idée de l’ampleur de l’exploit physique accompli : cet insecte de 40 mg et 10mm de long peut ainsi sauter à 30cm de haut, soit 25 fois la longueur de son corps et effectue jusqu’à six sauts périlleux avant d’atterrir … et tout ça avec un départ arrêté et sur le dos ! L’accélération impulsée par la détente atteint 380 fois la valeur de la gravité ; chez une autre espèce, le lacon ponctué, on cite même des valeurs de 700 fois ce qui serait parmi les records absolus connus chez des animaux sauteurs.
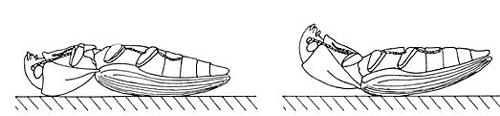
A aucun moment, ils ne déplient leurs ailes alors que ces insectes volent très bien. Les taupins peuvent sauter de la même manière en partant de la face ventrale mais avec une efficacité bien moindre. A noter que dans leur environnement, quand ils se retrouvent ainsi accidentellement sur le dos, ils essaient en principe d’abord de se relever avec leurs pattes. Par temps froid, placés sur le dos, ils font plutôt le mort et n’essaient pas de sauter ce qui suggère un coût énergétique important pour ce mode de retournement.
Mécanique du clic
Récemment, plusieurs équipes scientifiques se sont penchées sur le cas des taupins avec l’arrière-pensée de s’en servir pour la conception de robots articulés ; une équipe chinoise a d’ores et déjà réalisé un tel robot unijambiste de 8,5cm de haut capable de sauter quatre fois sa propre hauteur et utilisable dans des missions qui demandent d’entrer dans des espaces fermés à travers des passages étroits. Malgré toutes ces études approfondies, on n’a pas encore complètement percé tous les secrets de ce saut. Pour comprendre (un peu !), il faut se concentrer sur « l’organe du clic » situé sous la charnière entre le pronotum et les élytres, déjà mentionnée plusieurs fois, là où le corps se cambre puis se déplie ; sur un taupin vivant, il faut donc regarder l’insecte sur sa face ventrale juste au-dessus de l’insertion de la première paire de pattes. Là, on voit une pointe orientée vers l’arrière qui s’enclenche dans un creux du second article thoracique (mésothorax). Pour explorer le fonctionnement de cette structure, on a dû recourir au synchrotron à rayons X, notamment pour pouvoir suivre ce qui se passait à l’intérieur pendant la phase de projection en l’air.
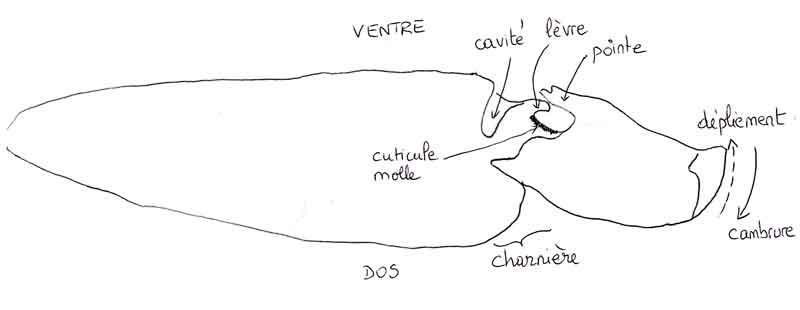
La cavité où peut venir se loger la pointe est surmontée d’une lèvre sur laquelle vient s’appuyer la pointe ; derrière cette lèvre, on note un creux correspondant à la charnière d’articulation et tapissé d’une cuticule souple particulière (la cuticule est la couche durcie qui recouvre l’épiderme des insectes). Ces éléments interviennent à tour de rôle lors des trois phases du saut.
Lors de la phase de pré-saut ou de mise en tension, le corps cambré vers le haut, la pointe glisse et vient se bloquer sur la lèvre comme sur un loquet. La cuticule souple se contracte et se déforme tandis que la pointe tourne lentement sur la lèvre sous la contrainte exercée par la contraction lente des muscles internes thoraciques. Cette phase qui dure donc plusieurs secondes permet le stockage d’énergie élastique comme dans un ressort.
A la fin de cette lente contraction, le verrou mis en place entre pointe et lèvre cède brutalement et la pointe glisse en avant pour aller se loger dans sa cavité d’accueil ce qui ramène le pronotum et la tête en avant ; ceci s’accompagne d’un rebond de la cuticule souple et la pointe oscille brièvement avant de venir se reposer dans sa loge. Quand la pointe atteint le fond de sa loge, la cuticule souple et la lèvre entrent en contact. Ce rebond élastique de la cuticule molle semble déterminant pour amplifier la force libérée brutalement avec de multiples oscillations en très peu de temps (dizaines de millisecondes).
On est donc sur le principe d’un mécanisme de ressort, connu chez divers autres insectes sauteurs, qui stocke de l’énergie élastique avant de la libérer brutalement en l’amplifiant ; mais la structure qui créé ce « ressort » reste elle unique et ultra-originale.
Déclinaisons
Ce système de ressort/loquet s’inscrit dans un ensemble de structures anatomiques très variées qui permettent aux petits animaux arthropodes (avec un exosquelette rigide articulé) d’accéder à des mouvements extraordinairement rapides et déployant une puissance étonnante. De tels mouvements ne peuvent être engendrés directement par la seule contraction des muscles : il faut à chaque fois un dispositif qui permette de stocker de l’énergie élastique sur un temps prolongé avant de la libérer brutalement en un temps très court.
Ainsi, chez les cicadelles (voir la chronique sur les cercopes) ou chez les sauterelles et criquets ou grillons (voir la chronique sur les orthoptères), le saut amplifié est permis par la contraction simultanée de deux muscles, un extenseur et un fléchisseur ; des articulations renforcées de forme particulière maintiennent le tibia (la jambe) en arc-boutant pendant cette phase lente de stockage d’énergie jusqu’à ce que le muscle fléchisseur soit inhibé ce qui déclenche une brusque détente du tibia et la projection en avant de l’animal. Chez les fourmis tropicales Odontomachus, ce sont les mandibules qui déploient une puissance incroyable en une fraction de seconde : un système de loquet ancré dans l’exosquelette de la tête les tient écartées l’une de l’autre ce qui stocke de l’énergie comme dans un ressort ; là aussi, un muscle déclencheur libère le verrou et les mandibules se referment en claquant à une vitesse folle (60 m/s), soit 2300 fois plus vite qu’un battement de cils. Elles se servent de ce « clac » soit pour tuer une proie, soit quand elles se retrouvent prises au piège au fond d’un entonnoir de fourmilion creusé dans le sable : là, elles font claquer leurs mandibules en les appuyant sur les parois ce qui les catapulte en l’air, hors du piège et hors de portée des mandibules redoutables de la larve de fourmilion ! Toutes proportions gardées, leur saut équivaudrait pour nous à un saut de 9 à 12 mètres. Et on pourrait ainsi multiplier les exemples pratiquement tous basés sur ce principe de base ressort/loquet mais avec à chaque fois des structures anatomiques très différentes.
Imparfait ?
D’un point de vue scientifique, on peut s’interroger sur la nature des pressions de sélection ayant permis le développement d’une telle structure avec un tel fonctionnement chez ces insectes ? Pour répondre, il faut d’abord observer un peu plus en détail l’efficacité réelle de ce saut au-delà des aspects purement mécaniques. Jusqu’ici, notre présentation donne l’impression de sauts « parfaits » et d’une efficacité redoutable. Sauf que si l’on suit statistiquement un certain nombre d’individus, on découvre que la probabilité de retomber sur le ventre est très proche de celle du hasard ; autrement dit, ce saut qui semble relever du coup de génie serait loin d’être vraiment efficace si le but est de se remettre d’aplomb. D’un point de vue physique, il suffirait que le taupin s’élève d’une fois la longueur de son corps et qu’il n’effectue qu’un demi-tour sur lui-même pour atterrir sur le ventre ; or, nous avons vu que le saut s’élève bien au-delà ce qui constitue une dépense d’énergie non négligeable même si le dispositif amplifie la puissance. Tout suggère qu’en fait, le taupin ne contrôle pas la puissance de son saut : elle est systématiquement maximale, bien au-delà de ce qui serait suffisant ; d’autre part, il ne contrôle pas non plus son orientation en l’air : chaque saut a sa propre combinaison imprévisible de roulements et de sauts périlleux !
Les modélisations indiquent qu’il lui suffirait de modifier de manière subtile ses points d’appui au sol quand il est cambré pour adapter sa vitesse, le nombre de rotations en l’air et l’angle de départ. Or, il saute toujours à pleine puissance et à la verticale. Donc, le taupin, faute de pouvoir contrôler sa belle mécanique du clic, saute le plus haut possible et compte sur la chance pour retomber à plat ! On est loin de l’outil « parfait »
Pourquoi ?

Avec ces données en tête, on peut maintenant aborder la question clé de la fonction réelle de ces sauts. De multiples hypothèses, dont certaines ne s’excluent pas, ont été émises depuis plus d’un siècle. Le bruit émis, le clic, pourrait servir à communiquer entre individus ou effrayer les prédateurs. Comme nombre d’espèces effectuent leur développement larvaire dans le bois mort, ce pourrait être un moyen au moment de l’éclosion de l’adulte depuis sa pupe de s’en extirper. On pense évidemment au moyen d’échapper aux prédateurs et aussi à cette possibilité de se remettre d’aplomb après une chute sur le dos. On peut essayer de « peser » ces hypothèses à l’aune des données accumulées.
L’hypothèse d’échapper aux prédateurs semble peu probable vu que l’insecte retombe presque sur place ; en fait, face à une situation d’attaque, les taupins font soit le mort en repliant pattes et antennes, soit ils s’enfuient. Le clic ne semble pas être un bruit suffisant pour faire peur : au plus, il peut surprendre une fraction de seconde ? Donc, on s’oriente plutôt vers un mécanisme spécifique pour se remettre à plat après une chute accidentelle sur le dos. Effectivement, la forme du corps et les courtes pattes ne facilitent pas la remise sur pied dans un tel cas.
Mais ce raisonnement « s’appuie » sur la situation où le taupin tombe sur le dos sur une surface dure ; or, dans leur environnement classique, ils vivent plutôt dans des milieux à végétation importante et se déplacent beaucoup sur les feuilles et les fleurs. Des chercheurs ont estimé chez une espèce de taupin que les surfaces des feuilles couvraient plus de la moitié de leur habitat. Or, quand un taupin tombe sur le dos sur une feuille et qu’il saute avec son clic, sa puissance de saut se réduit de … 75% du fait de la nature molle du substrat qui absorbe une part de l’énergie libérée. En même temps, cela abaisse leur vitesse de rotation sur eux-mêmes : ainsi, on se rapproche des conditions minimales évoquées ci-dessus qui suffiraient pour retomber à plat. Autrement dit, en déployant une puissance maximale à chaque saut, les taupins compensent, le plus souvent, la mollesse du substrat de départ !
Nature
Toutes ces considérations conduisent immanquablement à la question du « comment » tout cela s’est mis en place. Or, dans une des publications utilisées pour rédiger cette chronique, j’ai trouvé cette phrase (traduite de l’anglais) en guise de conclusion, phrase qui m’a laissé un peu pantois sachant qu’elle émane de scientifiques : « Que ces coléoptères utilisent une telle solution unique en son genre pour se remettre à plat est un exemple de plus de l’ingéniosité de la Nature à résoudre les problèmes ». Nature était bien écrit avec une majuscule dans le texte. J’y vois une perception divinisante de la nature qui s’appliquerait à résoudre les problèmes et qui aurait des intentions et aboutirait forcément à de géniales solutions. Il s’agit là de l’idée fausse classique qui consiste à utiliser la nature ou l’évolution comme sujet du verbe dans une phrase ; Guillaume Lecointre, dans la dernière version du Guide critique de l’évolution, développe avec sa grande clarté habituelle cette idée fausse (p. 162), une parmi les vingt-et-une qu’il a recensées. Je le cite donc sans retenue :

Les sciences visent à expliquer rationnellement le monde naturel. Si, en guise d’explication, l’acteur des phénomènes à expliquer se trouve être le monde naturel dans son ensemble, on n’a rien expliqué. … Mettre la nature comme sujet du verbe n’a aucune valeur explicative …
Dans ce cas des taupins, on peut juste dire que, au cours de l’évolution de cette lignée (évolution qui se poursuit toujours bien entendu), à un moment donné, par hasard (mutations), un premier changement dans l’articulation thoracique est apparu ; dans le contexte du moment (peut-être par exemple comme évoqué ci-dessus en facilitant l’extirpation de l’adulte de la pupe ?), cette modification a été sélectionnée positivement dans la mesure où elle procurait un avantage. Puis, d’autres modifications sont apparues toujours au hasard et elles ont été retenues ou pas selon différentes pressions de sélection. Il y a de fortes chances que cet exemple soit une exaptation, i.e. une transformation évolutive qui, au départ, avait été sélectionnée par rapport à une fonction donnée et qui, par la suite, s’est avérée devenir avantageuse pour autre chose (ici, se remettre à plat après une chute sur le dos ?). En tout cas, l’idée de l’intention ingénieuse de la Nature est battue en brèche par les imperfections nombreuses signalées ici : arrêtons de répéter que « la nature fait bien les choses » ; contentons-nous d’admirer la nature pour ce qu’elle est et c’est merveilleux !
Bibliographie
1)Coléoptères Elateridae de la faune de France continentale et de Corse. Leseigneur Lucien. In: Bulletin mensuel de la Société linnéenne de Lyon, 41e année, 1972. Coléoptères pp. 3-382;
Jumping without Using Legs: The Jump of the Click-Beetles (Elateridae) Is Morphologically Constrained. Ribak G, Weihs D (2011) PLoS ONE 6(6): e20871.
Latching of the click beetle (Coleoptera: Elateridae) thoracic hinge enabled by the morphology and mechanics of conformal structures Ophelia Bolmin et al. Journal of Experimental Biology (2019) 222,
Nonlinear elasticity and damping govern ultrafast dynamics in click beetles. Ophelia Bolmin et al. PNAS 2021 Vol. 118 No. 5
A Single-legged Robot Inspired by the Jumping Mechanism of Click Beetles and Its Hopping Dynamics Analysis. Gang Chen et al. J Bionic Eng 17 (2020) 1109–1125
The effect of natural substrates on jump height in click-beetles. Gal Ribak, Sonia Reingold and Daniel Weihs Functional Ecology 2012, 26, 493–499
