Le cou des oiseaux, précieux héritage dinosaurien
Aves

23/02/2022 Si on cherche les différences morphologiques majeures entre oiseaux et mammifères, on va en général citer en premier, et à juste titre, les ailes, le bec ou le plumage ; par contre, il n’est pas sûr que le cou vienne ensuite comme autre différence majeure. Et pourtant, il participe de manière déterminante à composer la silhouette unique des oiseaux. En plus, chez de nombreux oiseaux, le vrai cou est en grande partie caché par le plumage : il suffit de voir un squelette monté de passereau ou de pigeon par exemple pour réaliser qu’ils ont effectivement un cou bien plus long que celui des mammifères, à taille équivalente. D’aucuns vont opposer la girafe et son long cou mais il s’agit d’un animal de grande taille et elle est incapable de le replier : essayez d’imaginer une girafe se tordant le cou à la manière d’un flamant rose par exemple !

Flamants du Chili 
Le cou des oiseaux traduit sans doute le mieux l’étroite parenté des oiseaux et des dinosaures puisque les oiseaux ont émergé dans une lignée de dinosaures théropodes, la seule lignée de dinosaures qui ait échappé à l’extinction majeure de la fin du crétacé. Le cou des oiseaux a conservé en effet une bonne part des attributs du cou des petits dinosaures théropodes terrestres qui ont précédé les oiseaux comme par exemple le célèbre Velociraptor de Jurassic Park.

Bras de substitution
Quand on voit des oiseaux bouger leur cou dans leur vie quotidienne, nous nous sentons bien handicapés avec notre cou relativement rigide aux latitudes de mouvements limitées. En fait, le cou des oiseaux remplace le bras que les oiseaux n’ont plus dans la mesure où leurs membres antérieurs sont entièrement transformés en ailes dédiées entièrement au vol. Quant aux doigts devenus très réduits et inopérants pour des taches mécaniques, ils sont remplacés par le bec qui prolonge le … bras-cou !
Ce qui distingue le cou des oiseaux n’est pas tant sa longueur que sa flexibilité remarquable tout en conservant une grande force ; il contrôle la position et l’orientation de la tête qui porte les organes sensoriels et donc la main-bec. Il remplit ainsi une multitude de fonctions dont la plupart sont directement liées à deux traits majeurs des oiseaux : un corps couvert de plumes et le vol.

Le plumage demande en effet un entretien permanent et complet sur toute la surface du corps, queue et ailes comprises, pour conserver de bonnes capacités voilières, l’isolation thermique du corps et l’attrait visuel au moment de la reproduction. Se toiletter les plumes est une activité cruciale des oiseaux qui assure diverses fonctions : maintenir les barbes des plumes cohérentes entre elles (voir la chronique) en lissant les plumes avec le bec ; éliminer les débris d’usure et les nombreux parasites externes qui s’y installent dont les poux broyeurs de plumes ou mallophages ; récolter la sécrétion grasse de la glande uropygienne située sur le croupion et en enduire tout le plumage pour maintenir son bon état. Le cou long et flexible permet effectivement aux oiseaux d’accéder pratiquement à presque toutes les plumes de leur corps (sauf celles du dessus du cou mais certains y accèdent avec leurs pattes) et de les traiter avec leur bec.

Séance de toilettage chez un pélican frisé 

L’acquisition du vol s’est accompagnée de profondes transformations de tout le squelette bien au-delà des seules ailes avec notamment une extrême rigidification de l’ensemble thorax-abdomen et le renforcement de la cage thoracique ; les oiseaux ont un corps d’un « bloc » ! Or, en vol, il faut pouvoir contrôler ce qui se passe devant, derrière et sur les côtés en permanence surtout avec les vitesses souvent atteintes par nombre d’oiseaux : le long cou confère la mobilité de la tête permettant ce contrôle permanent d’autant que les yeux eux-mêmes sont très peu manœuvrables dans leurs orbites compte tenu de leur développement.

Thorax et abdomen forment un bloc soudé (poule) 
Héron cendré au décollage 
Héron cendré en vol, cou replié
Évidemment, le cou intervient aussi de manière décisive dans les activités de recherche de nourriture : localiser et suivre une proie tout en surveillant l’approche éventuelle de prédateurs ; atteindre le sol d’un coup de bec quand on a une haute stature ; fournir l’élan qui permet de frapper ou la force qui permet d’arracher ou de déchirer ; absorber les chocs quand on se sert du bec comme d’un marteau chez les pics ; … Enfin, de par sa mobilité, le cou intervient souvent lors des parades nuptiales en apportant la grâce et la complexité des mouvements.

Héron cendré prêt à frapper 
La détente du cou projette le bec en avant 
Spatule en train de barboter dans l’eau (photo R. Guillot)
Tenue et mouvements

Au repos, le cou est le plus souvent replié en S, calé dans le creux entre les deux clavicules. Cette forme globale laisse la liberté de mouvements au niveau de la tête et de la jonction avec le corps. Vu de l’extérieur, on se laisse très souvent berner par la longueur réelle car, ainsi replié, il est entouré du plumage du cou qui cache la courbure comme chez les canards. Les oiseaux à très long cou, comme les grues ou les cygnes, replient souvent complètement leur cou pour le reposer entre les ailes sur le dos ! En posture active, ils le tiennent vertical ce qui ramène le cou vers le centre de gravité du corps : ainsi, ces oiseaux n’ont pas besoin d’une musculature importante pour le maintien ni d’une queue importante qui contrebalance le cou.

Multiples postures du cou des cygnes 


En vol, la plupart des oiseaux tiennent le cou étiré, au moins pour ceux chez lesquels on peut le voir vraiment à cause du plumage. Mais certains oiseaux à long cou comme les hérons et apparentés ou les pélicans, le replient ; on pourrait invoquer l’encombrement d’un tel long bec mais d’autres oiseaux avec d’aussi grands becs comme les cigognes ou les grues volent le cou tendu.

Quatre positions du cou chez le canard colvert 


Pour bouger le cou, les oiseaux peuvent adopter trois grandes stratégies selon les groupes. Les poules par exemple qui se nourrissent au sol alors qu’elles sont haut perchées, doivent avancer la tête en avant et vers le bas : chaque vertèbre du cou pivote un peu quand le cou s’allonge mais aucune section du cou ne se replie ou déplie plus qu’une autre. Les cygnes ont un long cou mais des pattes relativement courtes : pour se nourrir, ils doivent allonger leur cou pour atteindre le fond de l’eau depuis la surface (voir la chronique « les girafes de la rivière »). Une vague de rotations s’enclenche en haut du cou et descend vers le bas jusqu’à la jonction avec le thorax ; au passage de cette onde, chaque vertèbre est tirée par la rotation de sa voisine jusqu’à obtenir un cou complètement étiré ; pour relever la tête, une vague inverse parcourt le cou et la pointe du bec est souvent la dernière partie à s’extraire de l’eau.

Cygne étirant son cou pour atteindre le fond de l’eau 
Enfin, chez les ratites (voir la chronique) comme les nandous, le cou est maintenant vertical tel un périscope ce qui permet de surveiller les environs dans leur habitat de steppes à hautes herbes. Pour se nourrir au sol, il doit abaisser la tête : une vague de rotations commence cette fois près du corps et remonte vers la tête ce qui l’abaisse à la manière d’un périscope. Elle peut remonter aussi vite de la même manière. Les grues utilisent aussi cette dernière technique. En pratique, on observe de nombreux intermédiaires entre ces trois techniques ou des combinaisons de deux d’entre elles selon la longueur du cou ou le mode d’alimentation.

Position basse du cou d’une autruche, « enroulé à l’envers » 
Cette remarquable mécanique toute en souplesse a inspiré des biomécaniciens dans la conception de robots avec un seul bras mobile construit sur le modèle du cou des oiseaux : des « ornithoïdes » en somme au lieu d’androïdes !

Deux postures du cou du héron cendré 
Colonne cervicale
Contrairement aux mammifères (voir ci-dessous) où la longueur du cou s’acquiert par allongement de chacune des sept vertèbres cervicales qui composent le cou, la longueur du cou des oiseaux varie via le nombre de vertèbres cervicales qui peut aller de (9) 11 à 25 ! Ainsi, les oies ont 17 à 19 vertèbres cervicales contre 16 chez les canards ou 24 chez les cygnes. On peut distinguer trois grandes zones le long de cette colonne cervicale délimitées par les points de flexion : le tiers supérieur capable de flexion ventrale, un tiers médian surtout dédié à la flexion dorsale et un tiers basal avec les deux types de mouvements mais de manière limitée. Chez le cygne, les 8 vertèbres en plus par rapport aux oies se situent dans la première zone.

Cou de poule ; noter les épines qui prolongent une partie des vertèbres cervicales 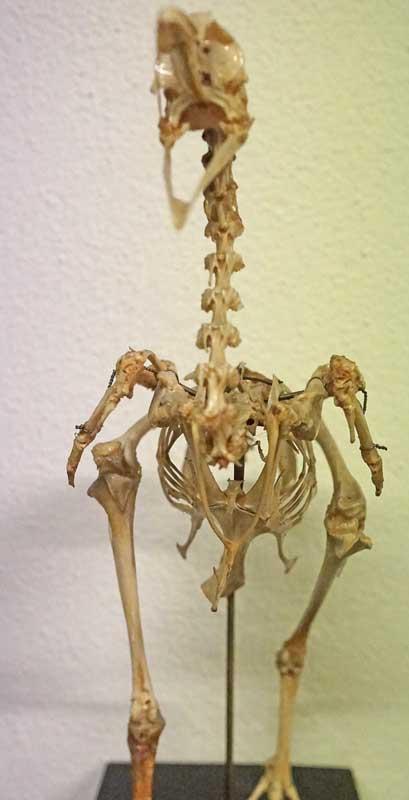

Dès les origines de la lignée des oiseaux au sein des dinosaures, les vertèbres sont caractérisées par des surfaces articulaires en forme de selle des deux côtés (hétérocoeles) : cette forme semble capitale car si les faces étaient plates, la colonne se déboiterait très facilement. De plus, la face en selle tournée vers la tête (crâniale) est sur le côté alors que celle de l’autre face se tient au bout si bien que ces faces décalées se bloquent entre elles à angle droit. De plus, cette alternance des deux selles orientées différemment aide à contrôler les mouvements aussi bien verticaux qu’horizontaux. Pour déformer son cou, un oiseau doit générer toute une série de petites rotations entre vertèbres.
Une partie des vertèbres porte des épines ainsi que des expansions dorsales qui limitent les risques de torsion, lesquels seraient très dommageables pour la moelle épinière et les vaisseaux sanguins qui passent dans ou sur cette colonne cervicale. A la jonction du cou avec le thorax, une paire de côtes cervicales (d’une seule pièce) vient renforcer la structure … comme chez les dinosaures théropodes.
Plus de 200 muscles, tendons et ligaments très complexes complètent cette colonne ; la plupart des muscles relient deux vertèbres non contigües ; un seul de ces muscles suit toute la longueur du cou. Ils forment des bandes musculaires très complexes que l’on retrouve quand on mange le cou d’un poulet : pas facile à décortiquer !
Double articulation
Au niveau de la jonction du cou avec la tête, on est frappé par la finesse de celui-ci par rapport au large crâne avec une impression de fragilité. Et pourtant, la solidité est bien au rendez-vous. Un seul condyle occipital (au lieu de deux chez les mammifères) sous le crâne entre en contact avec la première vertèbre cervicale ou atlas par une articulation en rotule, bien plus mobile que chez les mammifères ; ce condyle conserve la moelle épinière alignée avec le cerveau au-dessus quand elle s’engage dans le trou central (foramen magnum).
L’atlas, la première vertèbre cervicale, en forme de plaque fine, paraît elle aussi bien fragile. Elle est liée à la seconde vertèbre cervicale, l’axis, par du tissu connectif et surtout par un système de blocage unique en son genre : l’axis envoie un prolongement ou processus osseux (dens) qui s’engage dans une gouttière correspondante de l’atlas. Les deux vertèbres se retrouvent ainsi verrouillées ensemble et comme le processus osseux de l’axis émerge à la surface de l’atlas, l’articulation avec le crâne se fait donc via un double point de contact côté vertèbres, situation unique chez les vertébrés !

Fossile de Confuciusornis, un dinosaurien de la lignée basale des oiseaux 
Colonne cervicale d’un Dinosaure théropode (Tarbosaurus)
Comme indiqué dans l’introduction, ce type de cou si singulier est un héritage direct et bien conservé des dinosaures théropodes (voir la chronique générale sur les dinosaures) qui avaient eux aussi un long cou flexible mais disposaient encore de leurs membres antérieurs et de leurs doigts armés de griffes et avec un pouce. Dans la lignée divergente des Dinosaures sauropodes, on trouve aussi des cous gigantesques, démesurés. Ainsi le cou du brachiosaure Giraffatitan composé de 13 cervicales mesurait … 8,5m de long avec une seule vertèbre longue de 1,25m ! Pour Sauroposeidon, une estimation d’après des vertèbres partielles donne un cou de 11,5m de long ! Mais ce cou était basé sur une architecture très différente : en activité, il était maintenu horizontal, suspendu par des tendons et ligaments et la puissante musculature des épaules.
A noter cependant que le record absolu, si on raisonne en longue relative du cou par rapport au reste du corps, est détenu de loin par un animal fossile du groupe des prolacertiformes (une lignée proche de nos lézards actuels, ce qui n’est pas du tout le cas des dinosaures) : Tanystropheus du Trias, un animal probablement aquatique ; sur un spécimen long de 4,20m, le cou représentait 2,10m de long bien que ne comportant qu’une douzaine de vertèbres cervicales extrêmement allongées et fines.
Cas particuliers
Nous allons détailler trois cas particuliers d’oiseaux dont la structure du cou présente de fortes originalités.
Nous avons vu que la tête s’articule avec le cou de manière très simple et avec une certaine disproportion. Les mâles de certains Calaos, grands oiseaux des forêts tropicales, portent un bec énorme avec un casque frontal très dur ; l’ensemble représente presque 10% du poids de l’animal au bout de la tête perchée sur le cou ! Or, les mâles s’en servent pour s’affronter en joutes violentes où ils utilisent le casque comme une buttée ce qui génère un fort stress sur le cou. Chez ces oiseaux, l’atlas et l’axis sont complètement soudées et développent un second point d’articulation côté crâne en plus du condyle occipital (voir ci-dessus) : ceci limite les mouvements de côtés lors des affrontements. Même les espèces sans casque et à bec plus petit (mais quand même développé) ont cette structure unique.

Pélican frisé et son énorme poche sous le bec 
Anhinga africain et son long cou serpentiforme (photo R. Guillot)
Les cormorans, pélicans, fous et anhingas étendent leur cou sous l’eau pour capturer leurs proies ce qui impose une certaine résistance (ceux qui font de l’aquagym comprennent !) ; les fous plongent même depuis une grande hauteur en vol, le cou tendu, et pénètrent violemment ainsi dans l’eau ; les pélicans (voir la chronique) accumulent de l’eau dans leur poche gulaire avec les proies éventuelles et la relèvent comme une épuisette pour évacuer ensuite l’eau et avaler les poissons. Chez ces oiseaux, le cou a une structure renforcée comme une tour pétrolière ou derrick via de grosses vertèbres mais très fortement allégées (pneumatisées) qui s’empilent comme des soucoupes renversées se supportant les unes les autres.

Chez les chouettes, le plumage bouffant cache complètement la structure du cou
Les chouettes et hiboux (les strigiformes : voir la chronique) ont de très gros yeux immobiles ; chassant la nuit, ils doivent pouvoir écouter dans toutes les directions pour localiser finement leurs proies au bruit et ensuite fixer le point localisé avec les yeux. Pour ce faire, ils ont la capacité remarquable de pouvoir tourner la tête sur elle-même tout en gardant le corps immobile à … 270°. Imaginez un humain faisant de même : c’est la mort assurée car cette torsion sectionnerait la moelle épinière et cisaillerait les artères qui passent par la colonne cervicale. Plusieurs adaptations uniques permettent cet exploit : le canal central de la colonne cervicale est très grand ce qui laisse de la place à la moelle épinière lors des rotations ; les canaux par où sortent les artères sont dix fois plus grands que le diamètre de celles-ci au lieu d’être presque du même diamètre : ainsi, elles ne sont pas comprimées. De plus, les artères se dilatent en poches servant de réservoirs de sang contractiles qui assurent la libre circulation du sang même quand le cou est tourné : le cerveau conserve son irrigation sanguine. Enfin, il existe des anastomoses (des connexions) entre les artères vertébrales et les carotides : si une des deux voies est bloquée, l’autre peut prendre le relais.
Versus mammifères
A plusieurs reprises, nous avons souligné les extrêmes divergences entre le cou des oiseaux et celui des mammifères. La différence majeure tient avant tout au nombre de vertèbres cervicales : d’un nombre variable chez les oiseaux (mais aussi chez les lézards et serpents ou les amphibiens), elles sont toujours au nombre de sept chez tous les mammifères indépendamment de leur taille, même les girafes. En fait, les mammifères ont conservé strictement le nombre de vertèbres cervicales de leur lignée ancestrale qui remonte à plus de 200 millions d’années et a divergé de celle ayant ensuite donné les chéloniens (tortues), les lézards et serpents, les crocodiliens et les oiseaux. Ce nombre remarquablement constant au sein d’un groupe aussi diversifié dans ses modes de vie et dans sa morphologie a de quoi surprendre.

On a observé que, chez les mammifères, les anomalies individuelles du nombre de vertèbres cervicales (y compris chez l’espèce humaine) sont associées à un accroissement des risques de mortalité fœtale, de cancers très précoces et de problèmes nerveux ; de plus, ces risques s’accompagnent de divers problèmes physiques comme des nerfs, des muscles ou des vaisseaux sanguins déplacés ou comprimés. Ceci pointe vers des contraintes sur les gènes du développement qui agissent en même temps sur l’ensemble du développement du squelette et du système nerveux : toute mutation sur une partie de cet ensemble se répercute sur tout l’ensemble. Il se pourrait aussi que la susceptibilité aux cancers soit plus grande chez les mammifères que dans les autres groupes avec un nombre variable de vertèbres cervicales. On estime que le coût élevé de ces risques et de leurs conséquences a sélectionné contre ces mutations et conservé le nombre originel de sept vertèbres cervicales.

Il existe néanmoins deux exceptions à cette règle au sein des mammifères : les lamantins (siréniens) et les paresseux (Xénarthres). Chez ces derniers, les unaus (Choloepus) ou paresseux à deux doigts ont 5 à 7 vertèbres cervicales tandis que les bradypes ou paresseux à trois doigts en ont 8 à 9. Chez les unaus, il semble que ce soit la conversion de vertèbres cervicales en vertèbres thoraciques qui explique le raccourcissement du cou et inversement chez les bradypes. On retrouve de telles transformations chez des souris porteuses de mutations sur les gènes du développement. Le style de vie très lent et leur métabolisme global très bas les protègerait des effets délétères de telles transformations. Celles-ci seraient par ailleurs des adaptations au mode de vie arboricole de ces étranges animaux en leur permettant par exemple de tourner la tête bien plus que chez les autres mammifères quand ils sont suspendus à une branche.

Bibliographie
The inner bird. Anatomy and evolution. G. W. Kaiser. UBC Press 2007
Control of the Cranio-Cervical System During Feeding in Birds A. H. J. VAN DER LEEUW et al. AMER. ZOOL., 41:1352–1363 (2001)
Study on the Structural Characteristics of Bird Necks and Their Static Motion Features in the Sagittal Plane. Jiajia Wang et al. Coatings2021,11,1228.
Why sauropods had long necks; and why giraffes have short necks. Taylor and Wedel (2013), PeerJ 1:e36
Why Do Almost All Mammals Have Seven Cervical Vertebrae? Developmental Constraints, Hox Genes, and Cancer F. GALIS JOURNAL OF EXPERIMENTAL ZOOLOGY (MOL DEV EVOL) 285:1926 (1999)
Homeotic transformations reflect departure from the mammalian ‘rule of seven’ cervical vertebrae in sloths: inferences on the Hox code and morphological modularity of the mammalian neck Christine Böhmer et al. BMC Evolutionary Biology (2018) 18:84
