Le plumage des oiseaux, arc-en-ciel incomplet ?

11/08/2022 De tous les vertébrés terrestres, les oiseaux sont de loin ceux qui affichent la plus large palette de couleurs à nos yeux. J’ai collecté au hasard quelques noms vernaculaires évocateurs en feuilletant une encyclopédie des oiseaux du monde, histoire de rêver un peu : colibri topaze ; quetzal resplendissant ; lori féérique ; passerin arc-en-ciel ; jardinier ardent ; paradisier gorge-d’acier ; perce-fleur indigo ; tangara à col d’or ; brève grenadine ; manakin à tête d’opale ; …

Et encore, nous n’apprécions qu’une partie de cette incroyable palette de couleurs car les performances de notre vision des couleurs restent bien en deçà de celle des oiseaux eux-mêmes. Nous avons d’ailleurs consacré une chronique à la vision des couleurs chez les oiseaux et nous vous conseillons de la lire attentivement avant celle-ci car elle expose des notions de base que nous ne réexpliciterons pas ici. Pour autant, en dépit de cette diversité apparente qui nous subjugue, est-ce que cette palette de couleurs est aussi diversifiée qu’il nous semble ? N’y a-t-il pas des « zones d’ombre » dans cet arc-en-ciel, des couleurs non observables chez aucun oiseau ? Ont-ils réussi à dépasser en diversité de couleurs un autre groupe d’êtres vivants qui nous éblouit tout autant : les plantes à fleurs ? A toutes ces questions, un des grands spécialistes des plumes des oiseaux, R. Prum, et ses collègues apportent des éléments de réponse passionnants dans une publication clé de 2011. Pour comprendre, nous allons au préalable devoir nous frotter à certaines notions peu connues comme le gamut ou les espaces colorés ; mais l’enjeu en vaut vraiment la peine.

Gamut coloré avien
Jusqu’en 2011, en dépit de l’évidente diversité des couleurs des plumages des oiseaux et de la connaissance de leurs hautes capacités en matière de vision des couleurs, aucune étude n’avait cherché à cartographier l’ensemble des couleurs que peuvent renvoyer les plumages de tous les oiseaux, ce qu’on appelle le gamut coloré. En fait, à cette date, chez les vertébrés terrestres, on n’avait exploré cet aspect que chez les primates en recensant toutes les couleurs affichées par ces mammifères ; il est vrai que la tâche était bien plus facile du fait du nombre plus limité d’espèces (414 versus environ 10 000 oiseaux) et de la faible gamme de couleurs ternes de la majorité des pelages ; mais néanmoins, il y a de beaux signaux colorés comme les plages de peau nue du visage du mandrill. Il fallait donc impérativement effectuer cette étude sur les oiseaux et quantifier leur gamut coloré.

Le gamut coloré avien désigne donc l’ensemble des signaux visuels colorés émis par les différentes parties des plumages des oiseaux et perceptibles par les oiseaux eux-mêmes, i.e. des couleurs et tons avec des longueurs d’onde incluses dans leur spectre de vision qui, rappelons-le (voir la chronique) est plus large que le nôtre avec notamment un élargissement vers les UV.
Gamut est un mot d’origine anglo-saxonne, utilisé par la communauté scientifique sous sa forme francisée ; il a été créé dans le cadre musical où il désignait la gamme complète des tons disponibles pour une échelle musicale donnée (les sons, comme les couleurs, correspondent à des ondes). Comme au Moyen-âge, on transcrivait les notes par des lettres, la gamme sonore allait du sol le plus grave (alors noté avec la lettre grecque gamma) au do le plus aigu (noté ut) ; ainsi la gamme sonore s’écrivait gamma-ut ce qui a donné gamut. Donc, en dépit de l’homophonie partielle, gamut ne dérive pas de gamme.
Nous connaissons tous le gamut coloré des écrans d’ordinateur ou de téléviseur basé sur les 3 couleurs primaires ; ce peut être par exemple l’Adobe RGB très utilisé en impression ou le sRGB (standard) utilisé par défaut.
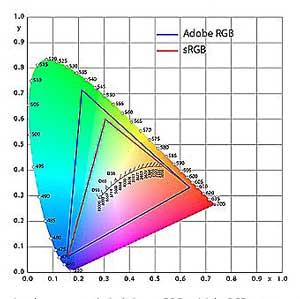
Espace coloré

Pour accéder à ce gamut avien, la première étape consiste à enregistrer le spectre coloré renvoyé par une tache colorée d’un plumage : une calotte, le dos, la gorge, la poitrine, le ventre, … Pour mesurer la lumière réfléchie ou réflectance, on éclaire, de manière standardisée, avec une fibre optique, une tache circulaire de 3mm de diamètre sur la partie échantillonnée et on enregistre les ondes renvoyées (entre 300 et 700nm pour être dans la gamme visible par les oiseaux) avec un spectrophotomètre. Ce travail s’effectue le plus souvent en laboratoire sur des peaux conservées dans les musées.

La seconde étape consiste maintenant à rendre ces informations lisibles en simulant comment seraient perçues les ondes renvoyées par ce « point coloré du plumage » à travers les yeux d’un oiseau, en vision tétrachromatique (voir la chronique sur la vision des couleurs). On calcule la stimulation relative des 4 types de cônes (L, M, S et UV) selon leur pic de sensibilité ; ces 4 valeurs sont reportées comme coordonnées dans un graphique en « 4D » sous la forme d’un tétraèdre dont les 4 sommets représentent les 4 types de cônes : ainsi chaque mesure d’une tache colorée d’une espèce apparait sous forme d’un point « suspendu » dans un espace en 4 dimensions. Ce graphique n’a pas la prétention de visualiser une expérience sensorielle qui dépend entièrement du traitement ultérieur effectué par les centres nerveux : il ne fait que cartographier dans un espace coloré la stimulation sensorielle sur les cônes d’une tache colorée.
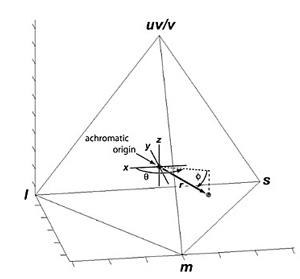
Les bords de ce tétraèdre représentent donc les frontières de toutes les couleurs imaginables qui sont théoriquement discriminables par l’œil d’un oiseau. Si l’ensemble des espèces oiseaux dispose effectivement de toutes les couleurs possibles, alors, avec un échantillonnage conséquent, on doit « remplir » tout l’espace interne du tétraèdre de points. Si par contre, il reste des zones de cet espace non occupées (aucun point), alors cela signifie que la couleur correspondante théoriquement possible n’existe pas chez les oiseaux ; il restera alors à expliquer pourquoi elle n’est pas disponible.
Échantillonnage
Comme il existe près de 10 000 espèces d’oiseaux, on comprend aisément qu’on ne peut pas (au moins dans l’immédiat) effectuer ce travail sur toutes les espèces. Il faut donc définir une stratégie d’échantillonnage qui permette de balayer de manière rationnelle et avec le moins de biais possible la grande diversité des oiseaux.

Les auteurs de cette étude ont analysé 965 taches colorées sur 111 espèces d’oiseaux différents, soit 1% de la diversité totale ; ce pourcentage peut sembler faible mais le protocole ci-dessous permet de compenser cet effectif. Selon les espèces, le nombre de taches colorées différentes peut aller de 1 (plumage très uniforme) à 7 pour les plus bigarrés. On choisit des espèces appartenant à des lignées différentes : ici, les 111 espèces se répartissent dans 55 familles classées dans 18 ordres. Ceci minimise considérablement la possibilité de rater un type de couleur particulier propre à une seule lignée. En plus, on a veillé à inclure le plus possible d’origines différentes des couleurs des taches.

Celles-ci peuvent être de deux types. Certaines proviennent de pigments très divers : mélanines, caroténoïdes (voir la chronique), porphyrines ou psittacofulvines. D’autres sont des couleurs dites structurelles ou physiques dues à la diffraction de la lumière par des nanostructures internes qui se répètent régulièrement comme au niveau des barbes ou barbules des plumes (voir la chronique) ; là aussi, on a veillé à inclure des espèces avec des couleurs structurelles générées selon des mécanismes différents. Ainsi, le diamant de Gould, bien connu comme oiseau de cage, qui figure dans cette analyse dispose de 6 couleurs différentes qui ont 4 origines différentes.

Gamut végétal
Les chercheurs ont élargi leur étude en cartographiant en parallèle le gamut de 2530 espèces de plantes dont 48 écorces, 590 feuilles et le reste en fleurs. Ces informations ont l’avantage d’être déjà disponibles dans une grosse base de données internationale, accessible à tous : Floral Reflectance Database. Ensuite, comme pour les plumages, ils ont converti ces mesures de réflectance végétale en points colorés dans l’espace coloré des oiseaux, i.e. comme si ces parties de végétaux étaient vues par des oiseaux. Mais pourquoi intégrer les végétaux dans une étude sur les couleurs des plumages des oiseaux ?
L’intérêt des végétaux est triple. Les oiseaux évoluent dans leur milieu au sein d’une végétation ; les signaux envoyés par leurs plumages sont donc en fait vus avec la végétation comme fond coloré ce qui peut modifier considérablement leur perception notamment via les contrastes. La comparaison des gamuts des oiseaux et des pantes à fleurs permettra de mieux comprendre comment les oiseaux ont accédé à leurs propres couleurs. Enfin, ce gamut végétal va servir de « contrôle » dans le sens où ces organes végétaux n’ont pas évolué en réponse à la vision avienne des couleurs vu que, à de rares exceptions près, ils ne sont pas ou peu consommés par les oiseaux : ils fournissent donc indirectement une estimation de la gamme des couleurs naturelles ne servant pas de signaux particuliers pour les oiseaux et qu’ils côtoient dans leur environnement. Certes, il existe des fleurs dites ornithophiles, i.e. visitées par des oiseaux nectarivores qui les visitent pour y récolter le nectar et d’ailleurs certaines d’entre elles ont été incluses dans les données. Mais, d’une part ceci ne concerne qu’un petit nombre d’espèces de plantes et on sait que leur coloration, souvent à dominante rouge, n’a pas évolué en fait sous la pression de la vision des oiseaux (plutôt pour échapper aux visites d’insectes). Par contre, on a exclu des organes végétaux testés les fruits car là, par contre, on a de nombreux cas de coévolution de la couleur des fruits en fonction du système visuel des oiseaux qui comptent de nombreux frugivores (voir la chronique sur l’endozoochorie).
Infinité limitée
Les résultats obtenus contredisent d’emblée notre idée reçue d’une infinité de couleurs chez les oiseaux : l’ensemble des points colorés des plumages testés n’occupe que … 26 à 30% de l’espace coloré total ; il y a donc bien moins de couleurs dans les plumages par rapport à toutes celles que pourraient percevoir les oiseaux. De plus, les couleurs des plumages ne sont pas uniformément réparties : il y a des « continents de teintes » où se concentrent de nombreux points séparés par des grands océans vides, sans aucun point.
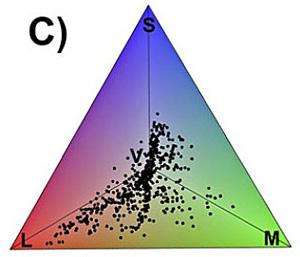
Les couleurs dues à des pigments chimiques n’apportent qu’une modeste contribution à cet espace coloré : seulement 7% du volume ; et chaque type de pigment n’occupe qu’un petit volume de l’espace : 3,5% pour les caroténoïdes, 2,7% pour les psittacofulvines, 1,5% pour les mélanines et 0,1% pour les porphyrines. Par contre, les couleurs structurelles représentent 17,9% du volume ; alors qu’elles ne concernaient qu’un quart des espèces testées, elles apportent 70% des couleurs du gamut total. L’acquisition de ce type de couleurs semble donc avoir été déterminante dans l’élargissement du gamut avien au cours de son évolution. On peut aussi penser qu’avec un échantillonnage plus grand incluant plus de telles couleurs, le gamut serait sans doute élargi et les 30% d’occupation sensiblement dépassés.
Les ultra-violets ne semblent pas apporter de couleurs en plus mais contribuent à une meilleure séparation des couleurs.
Versus végétaux
Qu’en est-il par rapport au gamut végétal ? Nouvelle surprise : les végétaux à fleurs occupent un espace coloré encore plus restreint : seulement 17% versus 26% pour les plumages. Donc, les oiseaux déploient incontestablement une plus grande diversité colorée que les plantes à fleurs … aux yeux des oiseaux qui sont dans cette affaire les observateurs. La rareté des couleurs structurelles chez les plantes explique une bonne part de cet écart.
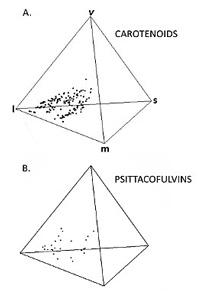
On constate un très fort chevauchement entre le gamut végétal et celui des plumages dans la gamme des rouge et jaune ; ceci n’a rien d’étonnant puisque les pigments responsables (voir la chronique), les caroténoïdes sont récupérés par les oiseaux dans leur alimentation végétale. La divergence entre les deux gamuts se fait dans les UV, les bleus et les verts. La majorité des couleurs structurelles des oiseaux tombent en dehors du gamut des végétaux : ceci suggère que l’évolution de la diversité de ces couleurs permettrait aux oiseaux de mieux se différencier de leur environnement naturel végétal.

Inversement, les plantes atteignent des couleurs absentes chez les oiseaux dans les pourpres et UV dues aux pigments anthocyanes uniquement végétaux : pélargonidine (rouge), cyanidine (magenta et bleu) et delphinidine (bleu). Ceci transparaît dans les noms vernaculaires : on a beaucoup de plantes « pourpres » (lamier pourpre, orchis pourpre, …) et pas d’oiseaux pourpres.
Syndrome de la rose bleue
Comment expliquer l’absence de certaines teintes dans le gamut avien comme le vert saturé, le bleu, les pourpres, les UV verts et UV rouge. Une piste d’explication repose sur l’hypothèse dite de la rose bleue. On sait que depuis longtemps les rosiéristes rêvent de produire une rose bleue ; or, c’est impossible car les roses ne fabriquent pas d’anthocyanes responsables de cette couleur chez les fleurs ; on y est arrivé en 2004 mais par génie génétique, i.e. en « trichant ».
L’examen détaillé des zones inoccupées dans l’espace coloré indique que les couleurs absentes seraient impossibles à produire avec les mécanismes de coloration existants. Chaque classe de pigment avien n’occupe qu’un tout petit volume de l’espace coloré et donc de nombreuses couleurs ne peuvent être atteintes avec les seuls pigments dont disposent les oiseaux. Ainsi, il n’y a pas de pigments bleus ; les oiseaux consomment des végétaux contenant des anthocyanes mais ils les digèrent entièrement sans avoir la capacité de les récupérer contrairement aux caroténoïdes. Les porphyrines vertes sont rares et ne donnent qu’une gamme restreinte de teintes.
Par contre, en combinant les couleurs structurelles avec les pigments, les oiseaux ont réussi à produire des couleurs saturées. Cette contribution majeure des couleurs structurelles confirme l’idée qu’elles ont évolué dans un contexte contraint par la limitation des pigments.
D’autres considérations complexes par rapport au fonctionnement des récepteurs suggèrent que nombre de couleurs saturées non atteintes sur les plumages des oiseaux seraient inopérantes car trop compliquées à percevoir.
Iles de Pâques

Les chercheurs ont été interpellés par au moins deux espèces dont le point coloré se retrouve complètement isolé au milieu d’une vaste zone inoccupée de l’espace coloré et qu’ils qualifient « d’îles de Pâques ». Les deux se situent dans la zone vide des pourpres mentionnée ci-dessus. Il s’agit de la calotte vive rose rouge du guêpier à fraise d’Asie et les rémiges primaires iridescentes du courol vouroudriou de Madagascar. Or, dans les deux cas, ces nouvelles couleurs proviennent de mécanismes spéciaux de production de couleur : pour le guêpier par la combinaison de caroténoïdes et de couleur structurelle et pour le courol par une couleur structurelle à partir des barbules des plumes. Il s’agit donc dans les deux cas d’exemples de sélection naturelle ou sexuelle liées à un environnement particulier et pas à une innovation majeure en soi.

(cliché Francesco Veronesi from Italy, CC BY-SA 2.0)
Innovations en séries
Justement, il est intéressant sur un arbre phylogénétique de pister où et quand sont apparues certaines innovations évolutives en matière de mécanisme de production de couleur. Ainsi, on découvre que les lignées basales (dont les Galliformes et ansériformes : voir la chronique) n’ont pas de caroténoïdes dans leurs plumages. La capacité à récupérer ces substances végétales n’est apparue qu’ultérieurement ; ensuite, de nouvelles innovations (équipement enzymatique) ont permis de récupérer les matériaux issus de la dégradation des xanthophylles (caroténoïdes jaunes) végétaux pour les recombiner en nouveaux caroténoïdes, des kétocaroténoïdes. On pourrait dire que les oiseaux ont réussi à s’affranchir en partie de leur dépendance aux végétaux pour ces couleurs et à les « faire leurs ».

De nombreuses autres innovations ont émergé dans diverses lignées, notamment au sein des passereaux, le groupe qui a connu une radiation évolutive considérable. Ainsi, le cotinga Pompadour (Amazonie) a réussi à développer un plumage pourpre bordeaux foncé (chez les mâles) via un mélange de six kétocaroténoïdes originaux.

De telles nuances bordeaux ne se retrouvent que chez les perroquets qui eux ont développé une nouvelle catégorie de pigments, les psittacofulvines ; ces pigments les placent à peu près dans le même espace coloré que celui des caroténoïdes des passereaux, le groupe frère des perroquets. Pourtant, les perroquets sont incapables de récupérer les caroténoïdes végétaux en dépit des quantités qui circulent dans leur sang et issus de leur alimentation ; ils ont étendu leur gamut en accédant à de nouveaux pigments. Ceci signifie que l’ancêtre commun aux perroquets et aux passereaux, avant leur divergence, n’avait pas non plus cette capacité ; les passereaux l’ont acquise au cours du développement de leur lignée.

Donc, globalement, les innovations évolutives liées à la production de couleurs ont souvent évolué séparément dans telle ou telle lignée et à de nombreuses reprises ; ceci est encore plus vrai pour le développement des couleurs structurelles au rôle majeur dans cette folle diversification.
Bibliographie
1)How colorful are birds? Evolution of the avian plumage color gamut Mary Caswell Stoddard and Richard O. Prum Behavioral Ecology Advance, 1-10 ; 2011
