Les pelotes des chouettes : des outils d’investigation

21/01/2021 Les rapaces nocturnes, comme les rapaces diurnes et divers autres groupes d’oiseaux (hérons, pie-grièches, martin-pêcheurs, guêpiers, goélands, …) rejettent par la bouche les restes non digestibles de leurs proies (os, poils, plumes, carapaces) sous forme de boulettes compactes, les pelotes de réjection. Le mécanisme de formation de ces pelotes a été particulièrement étudié chez les chouettes et hiboux et s’avère assez complexe. Ces pelotes présentent par ailleurs un avantage considérable par rapport à celles des rapaces diurnes notamment : une meilleure conservation des restes squelettiques des proies consommées. De ce fait, elles font l’objet de nombreuses études et analyses qui permettent de les utiliser comme de puissants outils d’investigation dans divers champs de la recherche scientifique.
Triple estomac
Les rapaces nocturnes ont tendance à avaler leurs proies entières dans la mesure où la taille de celles-ci est compatible avec l’ouverture du gosier ; pour les proies plus volumineuses, ils pratiquent une fragmentation minimale avec leur bec crochu en découpant la proie en gros morceaux avalés en entier à leur tour. La proie avalée descend rapidement le long de l’œsophage où, chez les chouettes et hiboux, il n’y pas de poche latérale de stockage temporaire de la nourriture, le jabot (voir la chronique sur le lait des pigeons) ; la proie atterrit dans la poche supérieure de l’estomac en trois parties, le proventricule, aux parois glandulaires qui sécrètent le suc gastrique. Rapidement, la nourriture s’engage vers la seconde poche aux parois musculeuses, le gésier. Le sphincter entre le proventricule et le gésier se ferme ; à l’autre extrémité, une troisième poche très réduite (quelques millimètres) et qui s’ouvre par-dessus le gésier, l’estomac pylorique, bloque le passage vers le début de l’intestin grêle. Sa fonction semble consister à filtrer le passage des éléments osseux ou des plumes susceptibles d’abîmer la fragile muqueuse intestinale. C’est dans le gésier que va se passer l’essentiel de la digestion et la formation de la pelote.
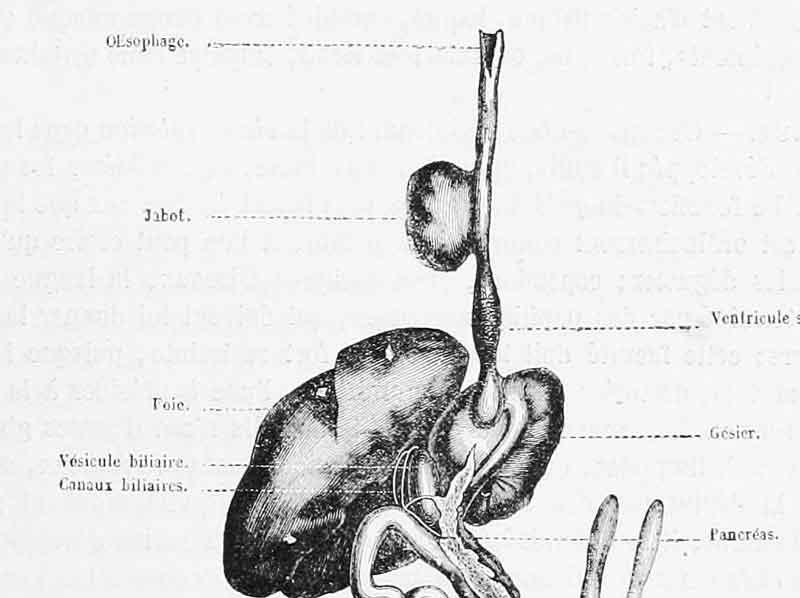
Plusieurs études utilisant notamment l’imagerie médicale ont permis de suivre ce qui se passait dans l’estomac de grand-duc de Virginie (espèce américaine). Le gésier se dilate fortement lors du remplissage avec des contractions de forte amplitude mais espacées ; ensuite, le gésier va progressivement diminuer de volume tout en restant rempli. Au cours des 4 à 8 heures qui suivent, la digestion chimique opère avec des contractions de faible amplitude et très espacées. Ensuite, pendant 1 à 2 heures, la chair liquéfiée est expulsée vers l’intestin grêle par des contractions de plus en plus rapprochées. Les éléments non digestibles (poils, plumes, os, carapaces, écailles, griffes, …) se retrouvent ainsi progressivement concentrés et compactés par les contractions du gésier en vase clos : la pelote se forme.
Digestion chimique
On aurait tendance à penser que les proies sont soumises à une puissante attaque acide dans l’estomac ; c’est effectivement le cas pour les rapaces diurnes où pH gastrique reste très bas autour de 1,6. Si on laisse des des os de souris dans une solution d’acide chlorhydrique de pH 1,6 à 40°C (t° corporelle des oiseaux), ils sont complètement dissous en une nuit. Et c’est bien ce qui se passe avec les rapaces diurnes dont les pelotes ne contiennent que très peu de résidus osseux sans que pour autant les os soient passés dans l’intestin. Par contre, pour les rapaces nocturnes, comme il a été dit en introduction, on observe dans la plupart des cas que les restes squelettiques sont relativement intacts : régulièrement, dans des pelotes d’effraie des clochers notamment, on arrive à trouver des squelettes quasi entiers de petits mammifères, y compris des os très fins comme les côtes.

Pelotes d’effraie des clochers 
Tibia de petit mammifère intact extrait d’une pelote 
Cubitus de petit mammifère intact extrait d’une pelote
Des études ont été menées sur des effraies avec des prélèvements gastriques réguliers par sonde suite à un repas. On constate alors que le pH varie considérablement au cours des huit heures du processus selon une gamme allant de 2 à 6. En fait, dès que la proie arrive dans l’estomac, le pH commence à monter (moins acide) et continue de le faire jusqu’à l’éjection de la pelote ; une heure après celle-ci, le pH chute brutalement à 2 avant de remonter et se stabiliser autour de 4. Il apparaît aussi que l’acidité serait plus forte (donc un pH plus bas) dans l’estomac des jeunes.

Contenu d’une pelote de Grand-duc : des restes d’insectes (Cerf-volant) et quelques débris osseux 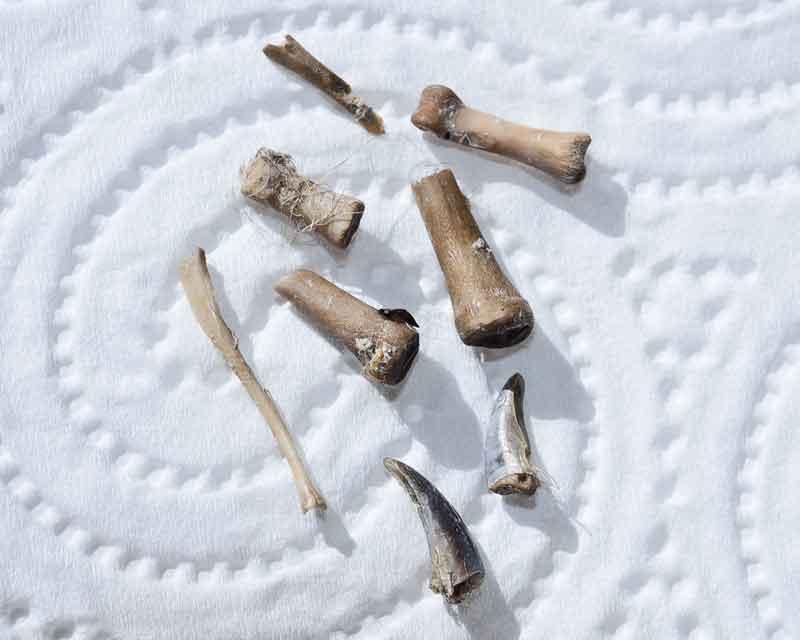
Détail des débris osseux très réduits, cassés, et griffes
A noter que la digestion chimique est assurée à la fois par le suc gastrique et par les sucs intestinal et pancréatique qui remontent via le pylore ; or, ces deux derniers n’agissent que sous un pH relativement neutre ; ils participent largement à la digestion des protéines ce qui accélèrerait le processus. Dernière particularité des chouettes et hiboux concernant cette fois l’intestin grêle : à la jonction avec le gros intestin viennent se raccorder deux tubes en cul-de-sac, des caeca (pluriel de caecum). Ils semblent jouer un rôle majeur dans la récupération de l’eau en recyclant les déchets azotés comme cela a été montré chez le grand-duc de Virginie. Le fait le plus curieux est que ces caeca sont absents par contre chez les rapaces diurnes sans que l’on sache expliquer la signification de cette différence.
Réjection
La réjection de la pelote se fait donc après la dissolution des chairs laissant os, poils et autres qui sont compactés par les contractions du gésier. Une douzaine de minutes en amont, des contractions de forte amplitude et très rapprochées (2/min) pendant 4 à 10 min (grand-duc) du gésier se déclenchent ce qui pousse la pelote dans le proventricule dont le sphincter s’ouvre ; ensuite, elle est poussée vers l’œsophage ou en quelques instants elle remonte grâce à des mouvements contraires au sens de progression (vers le bas) habituel, dit anti-péristaltiques et est rejetée par le bec.

Pelotes de grand-duc sur un rocher servant de reposoir 
Détail d’une pelote avec des plumes et un os cassé (probablement celui d’un oiseau)
Ce processus diffère du vomissement des mammifères carnivores qui ont un estomac simple ou de la rumination des ruminants ; dans ces deux cas, une seule contraction précède la remontée et non pas une longue série comme ici. Les mouvements anti-péristaltiques de l’œsophage s’observent dans la rumination mais pas pour le vomissement. Il s’agit donc là d’un processus original propre aux oiseaux dont les rapaces nocturnes.

L’intervalle entre la prise de nourriture varie selon les espèces de rapaces nocturnes, la taille des proies et le temps écoulé depuis le repas précédent ; logiquement, l’intervalle est plus court pour les espèces de chouettes et hiboux de petite taille et les petites proies (relativement à la taille du consommateur !). En captivité, chez un grand-duc préalablement mis à la diète, on observe un intervalle moyen de 8 heures et chez l’effraie de 6,5 heures. On observe souvent que le fait de présenter une nouvelle proie, sans même que le rapace la mange, suffit à déclencher la réjection sauf si cette présentation est trop rapprochée du repas précédent. En pratique, dans la nature, chez l’effraie par exemple, on trouve régulièrement les restes de 2 à 4 proies dans la même pelote. Mais quand les captures sont très espacées, le rapace peut très bien éjecter une pelote à partir d’une seule proie.
Débris squelettiques
Il semble clair que ce mécanisme évite à l’oiseau de faire passer tous ces débris non digestibles dans tout le tube digestif pour « rien » au bout du compte. La réjection libère l’estomac pour les repas suivants tout en allégeant le corps, un point essentiel pour la chasse. D’autre part, il évite les possibles blessures de l’intestin très fragile au passage de ces éléments durs et souvent pointus.
La comparaison de l’état des os dans les pelotes (fragmentation, cassures, dissolution partielle) permet de classer les différentes espèces selon l’intensité de destruction qui intéresse vivement les chercheurs désireux d’utiliser ces restes pour les « faire parler » (voir ci-dessous) ; on a ainsi défini cinq classes progressives de 1 à 5 allant de la moins destructrice (classe 1) pour l’effraie des clochers, à un peu plus (classe 2) pour le grand-duc du Cap à encore plus (classe 3) pour la chouette hulotte et le grand-duc européen. Dans les pelotes de ces derniers, les os petits et fragiles manquent souvent et un certain nombre d’os longs subissent une dégradation des têtes articulaires plus fragiles (cartilage). Il existe deux classes au-dessus mais elles ne concernent que les rapaces diurnes chez qui les os subissent une forte dégradation (voir ci-dessus la digestion chimique). L’observation des restes osseux au microscope électronique permet même de repérer une « signature » des rapaces nocturnes avec une forte densité de petites fissures courtes engendrées par la digestion chimique ; la taille de ces fissures augmente avec la taille des espèces.

Pelote de grand-duc riche en restes de coléoptères (lucanes) 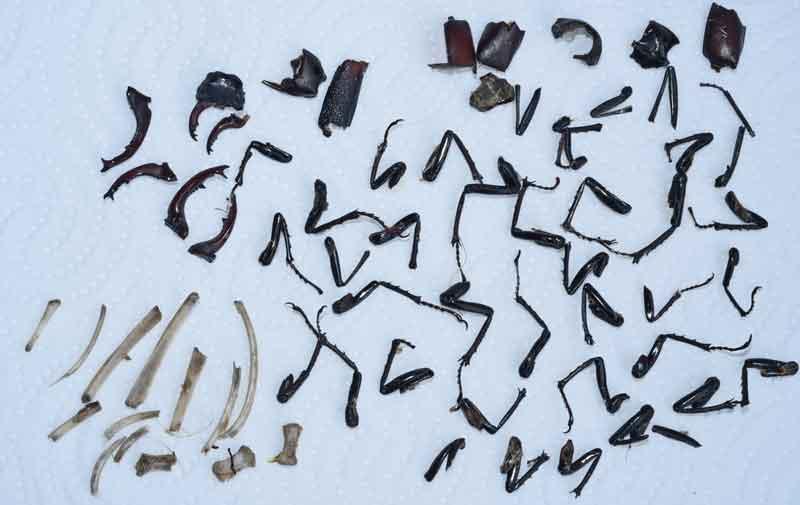
Analyse de cette pelote : pattes, mandibules, morceaux d’élytres (et quelques os fragmentés)
Le cas des carapaces d’insectes est plus complexe. Elles sont durcies par la présence d’une molécule résistante, un sucre polymérisé, la chitine. Des expériences en captivité ont été menées sur des hiboux moyen-duc nourris avec de la poudre de chitine ; environ 30% de la chitine absorbée se retrouve dans les pelotes recueillies ensuite où elle a conservé sa structure reconnaissable au microscope. Le reste a été digéré dans l’intestin grêle où sont secrétées des enzymes capables de la dégrader ; il est probable que les caeca mentionnés ci-dessus jouent un rôle dans cette digestion via des processus de fermentation bactérienne. Ces oiseaux en tirent donc un peu de ressources nutritives sous forme de sucres simples après dégradation. En temps normal, la chitine n’est pas ainsi fractionnée mais sous forme de pièces compactes et dures (élytres, pièces buccales, thorax) qui subissent moins d’attaque chimique et ne passent pas dans l’intestin, filtrées par le pylore, surtout quand elles sont pointues et potentiellement capables d’endommager la paroi intestinale.
Enquêtes écologiques

Cette préservation parfois presque totale des restes osseux comme dans le cas de l’effraie rend possible l’identification des espèces ingérées par une analyse très détaillée après dissection des pelotes en combinant les caractères des divers os (dont surtout ceux des crânes) et des dents pour les mammifères (voir site internet en bibliographie). Premier intérêt majeur : déterminer précisément le régime alimentaire de ces oiseaux, une tâche qui serait autrement fort délicate vu leurs mœurs nocturnes ! Ainsi des dizaines de milliers de pelotes ont été et sont toujours décortiquées par des ornithologues pour déterminer le régime alimentaire des différentes espèces et ses variations selon les habitats, les saisons, les pays, …

Mettre les pelotes à tremper 
Outils de dissection
Mais les pelotes présentent encore plus un intérêt considérable comme sources de documentation sur la composition des communautés de petits mammifères, les proies majeures de la plupart des espèces de rapaces nocturnes des régions tempérées : rongeurs et lagomorphes (lapins et lièvres qui ne sont pas des rongeurs : voir la chronique) et musaraignes (voir la chronique sur les Eulipotyphles). Or, ces petits mammifères représentent souvent une part essentielle de la faune des écosystèmes tempérés avec une très forte diversité d’espèces ; ils constituent par ailleurs des populations très nombreuses avec des taux de reproduction élevés, susceptibles de réagir très vite aux moindres changements environnementaux (habitats, climat, perturbations humaines). Et pourtant, leur étude s’avère très compliquée par leur petite taille, leurs mœurs nocturnes et leur vie cachée qui rend les observations directes très limitées. La seule solution scientifique repose sur le piégeage avec des appâts. Mais la mise en place de ce procédé soulève des problèmes techniques considérables dont le coût et le côté très chronophage sans oublier les aspects éthiques (capturer des animaux). En pratique, on ne peut guère piéger que sur des surfaces restreintes dans un seul habitat et sur une période temps limitée : très difficile d’extrapoler ensuite à des échelles plus grandes et dans des mosaïques de milieux. Si on n’utilise qu’un seul type de piège et d’appât, on introduit un biais procédural majeur en ne collectant qu’une partie des espèces présentes.

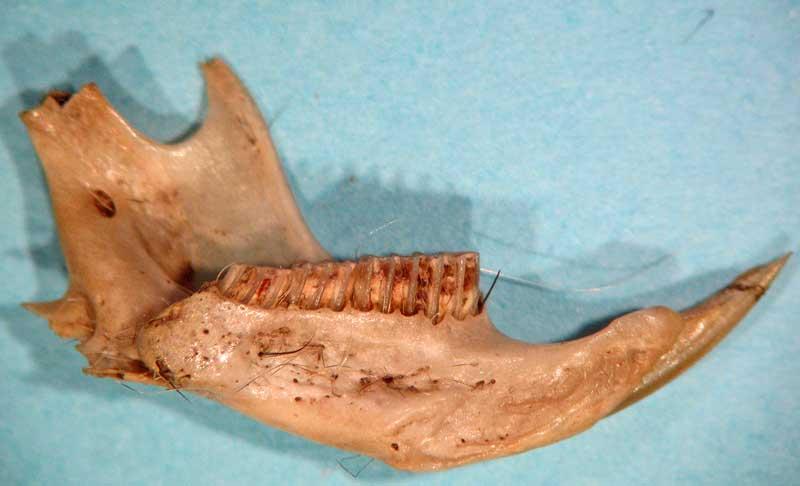
Le contenu d’une pelote d’effraie : en haut les os des membres ; en bas les crânes et mandibules 
Crânes et mandibules de musaraignes (dents de « carnivores ») 
Crânes et mandibules de petits rongeurs : mulot (en haut à droite) et campagnols (les plus courants)
Au vu de ces difficultés, il apparaît que l’analyse des pelotes de rapaces nocturnes récupérées dans les habitats étudiés constitue une solution puissante, bien moins onéreuse et surtout bien plus efficace en termes d’échelles de temps et d’espace. Chouettes et hiboux chassent sur de vastes territoires : 680 hectares en moyenne pour une effraie, 2500 hectares pour un grand-duc ou 70 hectares pour une chouette hulotte. La plupart sont des prédateurs plutôt généralistes qui recherchent une large gamme de proies et chassent dans divers habitats. Leur sédentarité facilite la tâche et rend possible un suivi sur l’année selon les saisons. Certes, il existe aussi quelques biais dans cette méthode comme la tendance à la sélection de certaines proies selon les abondances, la disponibilité et la visibilité. En contrepartie, on accède ainsi aux espèces rares souvent ratées dans les piégeages. Enfin, certaines chouettes comme l’effraie ont des répartitions mondiales très vastes ce qui autorise des comparaisons à très grande échelle spatiale quant à la composition des communautés de petits mammifères.
Mandibule de campagnol 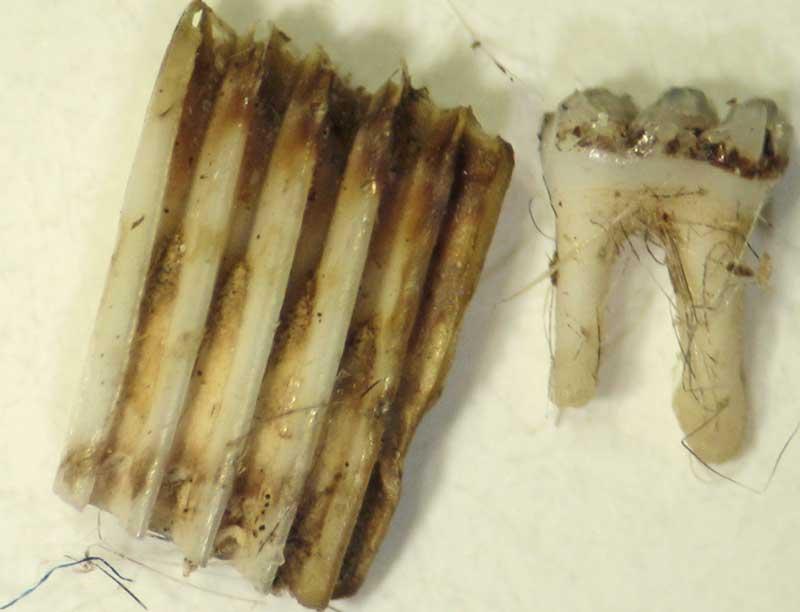
Molaire de campagnol (à gauche) et de mulot (à droite) 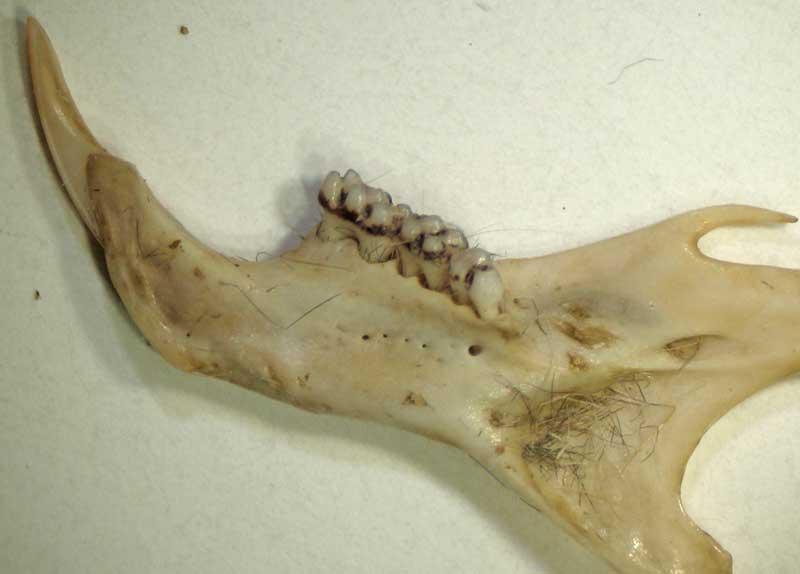
Mandibule de mulot 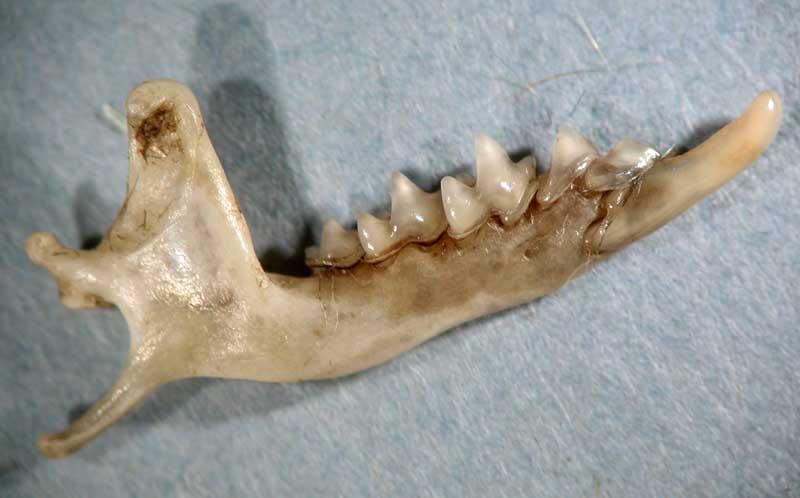
Mandibule de musaraigne à dents blanches (genres Crocidura/Suncus) 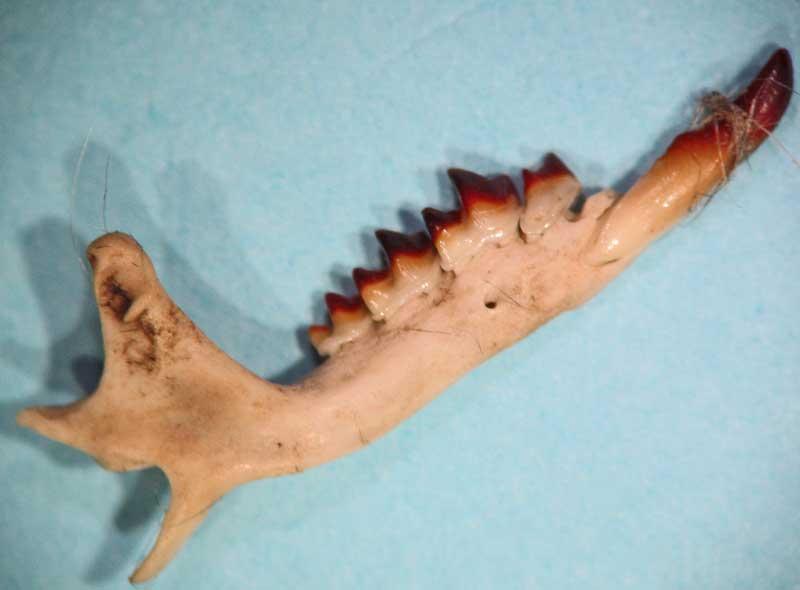
Mandibule de musaraigne à « dents rouges » (genres Neomys/Sorex)
Enquêtes archéologiques
Les rapaces nocturnes offrent un autre avantage : leur grande fidélité tout au long de l’année à des reposoirs, des sites où ils viennent se percher pour passer la journée, se reposer le jour et où ils déposent jour après jour leurs pelotes qui s’accumulent au sol. Or, certaines espèces dont le grand-duc d’Europe ou l’effraie des clochers habitaient autrefois (et continuent actuellement outre de nouveaux habitats créés par l’homme) souvent des sites rocheux du type cavernes, abris, falaises, … lesquels étaient justement aussi des sites recherchés et habités par les humains au Néolithique (voir la chronique La pierre est en nous). A l’occasion de fouilles archéologiques sur ces sites, on retrouve donc souvent des accumulations de restes osseux de petits mammifères et d’oiseaux dont une bonne part provient de l’activité de rapaces nocturnes (mais aussi de petits mammifères carnivores). Contrairement à une idée reçue, on s’est rendu compte que ces petits animaux pouvaient aussi être été chassés par les hommes au même titre que les plus grands. Ceci pose un premier problème : celui de déterminer devant des restes osseux s’ils résultent de l’activité prédatrice de rapaces (et si oui lesquels) ou de celle d’humains, objet de recherche des fouilles. Les signatures évoquées ci-dessus (et celles connues laissées par les hommes) permettent de faire la part des uns et des autres ; ainsi, on a constaté que les hommes qui occupaient la région de Gibraltar capturaient beaucoup d’oiseaux dont de grandes espèces, y compris les rapaces nichant à proximité (voir la chronique).

Accumulation de pelotes d’effraie dans un moulin en ruine servant de reposoir 
Le grand-duc capture des hérissons mais il arrache la peau avant de les consommer ! 
Pelote de grand-duc au sol et en cours de désagrégation : des os vont être perdus.
Mais ces accumulations « fossiles » de pelotes de rapaces nocturnes offrent un second intérêt majeur : reconstituer les paysages écologiques environnants dont leur faune en petits mammifères et leur évolution sur des pas de temps assez courts de l’ordre du millénaire ; il faut pour cela être d’abord capable d’identifier l’auteur des pelotes. Ensuite, pour exploiter la richesse considérable potentielle d’informations portées par ces restes, il faut conduire des études actuelles sur les modalités d’accumulation de ces pelotes, leur évolution, leur contenu afin de transférer ces données dans le passé ; c’est le principe de la taphonomie (de taphos, enfouissement), la science qui étudie la formation des gisements fossiles et tous les processus de transformations des objets naturels fossilisés. Ainsi, on a soulevé un certain nombre de problèmes quant à la représentativité réelle de ces vestiges parfois en grandes quantités. Selon l’espèce de rapace, la gamme des proies préférentielles ne sera pas la même. Se pose le problème des grosses proies préalablement « traitées » (décapitées, démembrées, fragmentées) et dont les restes peuvent de ce fait se retrouver dans des pelotes différentes. Selon l’altération des os conservés au fil des temps, les espèces proies les plus fragiles vont être désavantagées. Une fois au sol, les pelotes subissent aussi des transformations : consommation par d’autres animaux, déplacements, fragmentation, enfouissement, … Dans les pelotes fragmentées, les poils se dégradent et ne protègent plus les os ; ainsi sur des pelotes de grand-duc tombées à terre on passe de 99% des os intacts dans les pelotes fraîches à moins de 75% dans les pelotes fragmentées.
Enquêtes génétiques
Enfin, plus récemment, on a exploré un autre usage des pelotes de rapaces nocturnes comme source d’ADN pour la génétique des petites espèces de mammifères. Nous avons vu les fortes limites du piégeage pour la connaissance de la composition en espèces des communautés de petits mammifères : elles valent tout autant pour la connaissance de la diversité génétique au sein de chaque espèce selon ses populations sur l’ensemble de l’aire de répartition. D’où l’idée d’utiliser les os et les dents récoltés dans des pelotes comme source d’ADN permettant de réaliser de telles études. Une étude portant sur les os de 62 individus différents de petits rongeurs ingérés par des chouettes ou hiboux montre une forte variabilité de la préservation de l’ADN ; celle-ci est forte entre os d’une même pelote ce qui signifie que lors d’études génétiques, il faut tester les restes de plusieurs individus d’une même pelote. Par contre, la nature des os (crâne ou membres par exemple) et leur observation microscopique ne dit rien sur le degré de préservation de l’ADN. Même des pelotes récupérées dans des collections de muséums peuvent encore fournir de l’ADN exploitable. Une étude polonaise a exploré le cas d’un petit rongeur rare et localisé, le rat des moissons, difficile à échantillonner : en étudiant les mandibules trouvées dans des pelotes, ils on trouvé cinq fois plus d’ADN dans les dents que dans les os des mandibules. La qualité de l’ADN et la possibilité de l’amplifier dépendent de l’âge relatif des pelotes ; les chercheurs notent néanmoins une certaine dégradation de l’ADN. Cette ressource pourrait permettre entre autres de rechercher des espèces cryptiques, i.e. très proches morphologiquement mais bien distinctes génétiquement.
Bibliographie
Site LPO AURA sur l’identification des petits mammifères à partir des crânes trouvés dans les pelotes
Gastrointestinal physiology and nutrition in wild birds. GARY E. DUKE Proceedings of the Nutrition Society (1997), 56, 1049-1056
Owls, multirejection and completeness of prey remains: implications for small mammal taphonomy. Frédéric LAUDET, Christiane DENYS and Frank SENEGAS. Acta zoologica cracoviensia, 45(special issue): 341-355, 2002
Owl Pellet Taphonomy: A Preliminary Study of the Post-Regurgitation Taphonomic History of Pellets in a Temperate Forest. REBECCA C. TERRY. PALAIOS, 2004, V. 19, p. 497–506
Prey size as a critical factor for bird bone taphonomy in eagle owl (Bubo bubo) pellets. Anna Rufà & Véronique Laroulandie Scientific RepoRtS (2019) 9:19200
Owl pellets: a more effective alternative to conventional trapping for broad-scale studies of small mammal communities. Leanne M. Heisler, Christopher M. Somers and Ray G. Poulin. Methods in Ecology and Evolution 2016, 7, 96–103
High efficiency protocol of DNA extraction from Micromys minutus mandibles from owl pellets: a tool for molecular research of cryptic mammal species. Magdalena M. Buś et al. Acta Theriol (2014) 59:99–109
Owl pellets: a wise DNA source for small mammal genetics. S. Guimaraes et al. Journal of Zoology (2015)
