Une expérience grandeur nature qui décoiffe

La fragmentation des habitats, i.e. la transformation d’un milieu continu en de nombreuses taches séparées, plus ou moins grandes et isolées les unes des autres, entraîne des conséquences importantes sur les capacités de dispersion des espèces selon leur mobilité ou la nature de leurs agents de transport. Si la dispersion devient très limitée (par exemple si les fragments se trouvent très loin des autres dans une matrice par ailleurs inhospitalière : voir la chronique sur le cas de la porcelle enracinée), alors la persistance de l’espèce peut devenir problématique faute d’échanges de gènes avec des risques de consanguinité croissante et l’impossibilité de recolonisation en cas d’extinction dans un fragment. On sait que dans de tels cas l’existence de corridors linéaires permettant à minima la dispersion des espèces joue un rôle positif considérable pour contrer les effets ci-dessus. Si on a beaucoup étudié l’impact des corridors dans les paysages fragmentés sur les animaux ou sur les végétaux transportés par ces derniers (épi et endozoochorie), on ne sait que très peu de chose en ce qui concerne les organismes dispersés par le vent dont les végétaux dits anémochores, i.e. dont les graines/fruits sont déplacés par le vent (voir l’ensemble des chroniques autour de ce thème). En fait, il s’avère très difficile de suivre le devenir de telles graines dans des paysages très fragmentés et une équipe américaine a conçu une expérience incroyable tant dans l’espace (grandeur nature) que dans le temps (sur douze ans) ; cette chronique, au delà des faits scientifiques démontrés, s’attache d’ailleurs à illustrer l’ingéniosité des chercheurs en général et leur capacité à inventer des protocoles vraiment originaux et d’une incroyable minutie !

Fermeture

L’étude se situe en Caroline du sud sur le site de Savannah River, près d’un centre de traitement des déchets nucléaires, où une pollution au tritium, un radionucléide de l’hydrogène, a conduit à la plantation massive de forêts de pins censés évacuer le tritium par évapotranspiration ou le fixer dans le milieu. Ces plantations à grande échelle (sur plus de 1000 hectares) ont réduit l’écosytème originel, une savane herbacée à pin des marais très sujette à des incendies répétés, un milieu très ouvert, à l’état de fragments ou taches dispersées donc dans une matrice boisée et dense. Or, ce milieu abrite une riche flore très originale dont la majorité des espèces se dispersent par le vent (anémochorie). On se retrouve donc dans un contexte équivalent à celui que nous connaissons dans de nombreuses régions de France en forte déprise agricole où la reconquête naturelle par certaines essences (voir l’exemple des pins dans la Chaîne des Puys ou dans les Causses) ou suite à d’innombrables plantations (en timbres postes comme on dit) d’épicéas ou de sapins de Douglas : dans ces paysages devenus à dominante forestière, des milieux ouverts tels que pelouses, prairies ou zones humides se retrouvent comme des ilots perdus dans un océan d’arbres.

Notons à ce propos que l’on vante souvent en France le fait que la forêt gagne du terrain comme argument en faveur de l’environnement (dont une partie via des plantations) mais on oublie de dire que cela engendre des conséquences négatives sur la biodiversité des milieux naturels ouverts qui se retrouvent enclavés !

D. L. D.
La fragmentation des milieux ouverts par la reforestation agit sur le vent de deux manières : la canopée créé un obstacle vertical majeur à la propagation des graines/fruits emportés par le vent et les effets de lisière modifient la circulation et la vitesse des vents. Par rapport au premier impact, le maintien ou la création de corridors linéaires adéquats constitue donc une réponse permettant la circulation des graines/fruits anémochores (mais aussi celles transportées par les animaux !) entre fragments de milieux isolés. Il ne faut pas non plus oublier à cet égard l’importance majeure de la circulation des pollinisateurs qui utilisent aussi ces corridors mais cet aspect n’est pas le sujet étudié ici. Le second effet s’avère plus complexe et dépend entre autres de la forme des fragments.
Concernant la dispersion, il faut distinguer deux composantes aux effets bien différents par rapport à la problématique de la fragmentation. La dispersion à courte distance reste prédominante : les graines/fruits se décrochent et soit tombent au pied par temps très calme, soit se déplacent de quelques mètres à dizaines de mètres emportées par le vent. Les études montrent qu’une majorité écrasante de graines/fruits anémochores suit ce processus et atterrit dans un rayon limité par rapport au pied mère. Cette composante permet le maintien de l’espèce sur place, in situ dans son fragment de milieu, notamment pour les espèces annuelles qui doivent se renouveler chaque année. Mais on reste « entre soi », entre membres d’une même population et à moyen terme le risque de consanguinité, et donc de perte de variabilité essentielle pour s’adapter aux changements, prend forme et grève les chances de maintien de cette population.
Pour que des fragments éloignés puissent échanger entre eux, il faut qu’aient lieu des évènements de dispersion à longue distance (D. L. D.), i.e. des graines/fruits transportés exceptionnellement sur de longues distances pouvant excéder le kilomètre et plus. Démontrer l’existence de ces DLD s’avère très difficile sur le terrain mais on sait qu’ils existent même s’ils ne concernent souvent que moins de 1% des graines/fruits libérées par une population donnée ! Un seul de ces évènements DLD peut suffire à apporter du « sang neuf » dans un fragment longtemps resté isolé. Or, pour que ces DLD puissent avoir lieu pour des graines anémochores, il faut des conditions particulières réunies : des épisodes de vents violents en période de dispersion (fin automne/hiver) et sur place des conditions physiques qui permettent la création de courants d’air ascendants suffisamment puissants pour franchir la fameuse barrière des arbres en lisière. Autrement dit, pour étudier ce problème, il faut s’intéresser à la dynamique des vents en fonction de la topographie, des effets de lisières et de la présence des corridors : l’essentiel de cette étude et la conception du protocole minutieux sont basés sur cette connaissance.
Création sur mesures
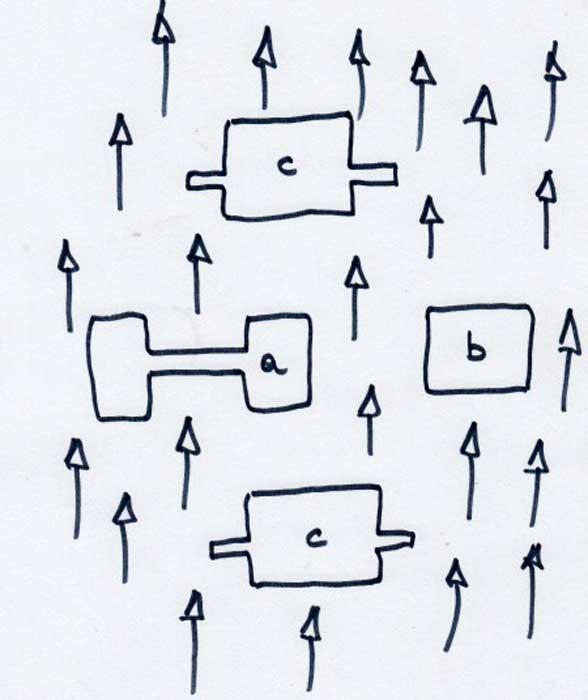
Dans une matrice de forêt plantée de pins des marais (Pinus longifolia) et de pins à encens (P. taeda), haute de 22m en moyenne, l’équipe de chercheurs a délimité des fragments ou taches sous forme de clairières alignées en parallèle : une gestion adaptée a permis la reconstitution du milieu originel, une savane claire ouverte (voir ci-dessus) ; il y a donc un très fort contraste entre ces taches herbacées ouvertes et la forêt haute et dense avec des lisières « abruptes ». Sur un plan schématique ci-joint, réalisé d’après la photo aérienne des parcelles recréées, nous avons reporté les trois types de taches légendées a, b et c. La tache (a) est formée de deux taches de 100m sur 100m connectées entre elles par un corridor de 150m de long sur 25m de large ; b et c sont des taches non connectées : (b) est une tache rectangulaire de 100m sur 134,5m et (c) des taches « ailées » de 100m sur 100m prolongées sur deux côtés par deux corridors (en forme d’ailes vues du ciel !) de 75m sur 25m mais non connectés avec une autre tache. La précision des mesures vise à s’assurer que toutes les parcelles ont exactement la même surface (avec ou sans corridor) de manière à permettre les comparaisons. Ces parcelles expérimentales sont suivies depuis douze ans au niveau de leur flore, bien plus riche en espèces que celle des forêts plantées environnantes !
Pour suivre la dynamique du vent (voir ci-dessus) des tours météo locales ont donné les vitesses de vent au-dessus de la canopée ; des anémomètres placés à différentes hauteurs dans les différentes taches relèvent en continu vitesses et directions des vents lors des périodes de dispersion des graines (septembre à décembre ici). Avec ces données et un modèle météorologique préexistant, les chercheurs ont ainsi pu élaborer un modèle ultra-précis (intégrant le volume en 3D des feuillages) et surtout le tester ensuite en le confrontant avec les données de terrain. Ce dernier point est majeur car, jusqu’ici, les études sur l’anémochorie et les évènements DLD se basaient sur des modèles théoriques pratiquement jamais ou très peu testés ensuite sur le terrain.
Lâcher de graines

Cirse d’Angleterre dans une prairie humide 
Fruits plumeux transportés par le vent 
Arnicas dans une lande montagnarde enclavée 
Fruits à aigrette plumeuse des arnicas
Reste maintenant l’élément clé de l’histoire : les graines/fruits des espèces anémochores de ce milieu ouvert. La connaissance de la flore locale a permis de déterminer les types de graines/fleurs et la vitesse terminale de ces graines lors de leur chute : plus la graine chute lentement (en étant freinée par exemple par des structures en parachutes de poils soyeux : voir la chronique sur le salsifis douteux), plus elle aura de chances d’être prise en charge par un courant d’air et déplacée, voire soulevée en cas de courant ascendant. Impossible de suivre individuellement des graines/fruits surtout avec plusieurs espèces : les chercheurs ont donc opté pour une solution originale, créer des graines artificielles faites de fibres synthétiques de sorte qu’elles entrent dans la gamme des vitesses terminales des graines/fruits locaux. Là encore, le protocole suivi épate par sa précision et sa rigueur : pour chaque tache (a, b et c), on lâche depuis une hauteur de 4,50m, à raison de cinq graines toutes les 30 secondes pendant 30 minutes, en sept endroits sélectionnés de la tache et ce pendant cinq périodes avec une direction unique de vent pendant le lâcher. Au total, 5400 de ces graines artificielles ont été ainsi libérées ; des teintures de couleur différente permettent de savoir précisément d’où ces graines ont été relâchées. Ensuite, grâce à leur haute détectabilité (dont la teinture colorée), les graines sont recherchées et géolocalisées dans les taches ! Au total, dix de ces taches étaient réparties sur une surface de 50 hectares, soit une expérience vraiment grandeur nature et inédite !

Marais à angélique et molinie entouré de plantations de résineux 
Fruits ailés anémochores des angéliques
Triple effet
Il en résulte des milliers de données à analyser et interpréter avec, tout au bout, enfin des résultats ! Trois effets distincts de ces ouvertures dans une forêt par ailleurs fermée sur la dynamique du vent ont pu être mises en évidence. Toutes les taches créent un effet de réorientation du vent autour de leur axe le plus long, en alignement avec le corridor ou les ailes en notant bien que l’orientation des taches et de leurs « appendices » n’avait pas été faite selon la direction des vents dominants. Cet effet spatial est le plus marqué dans les taches à corridor, intermédiaire dans les taches ailées , et minimal dans les taches simples ; l’effet est maximal près du sol. Par conséquent, on voit que les graines/fruits seront donc déplacées plus vite dans les taches à corridor et orientées de plus dans le sens de ce dernier.
Quand le vent accélère, il subit un second effet de soufflet qui conduit à une intensité plus grande à l’extrémité des corridors et des ailes sous le vent ; l’augmentation de la vitesse est plus forte au centre des ailes ou des corridors ; les graines/fruits vont donc être poussées vers le corridor encore plus fort ; par contre, pour les taches ailées, les graines vont atterrir dans la matrice boisée ! Enfin, les résultats mettent en évidence des « points chauds d’éjection », i.e. des zones ponctuelles dans les taches où la conjugaison de l’accélération, de la réorientation et de l’apparition de turbulences avec une forte composante verticale induit une ascension de la colonne d’air propice à un événement de DLD.
Finalement, il se confirme que les taches connectées par des corridors linéaires ont beaucoup plus de chances de voir se produire des évènements de DLD susceptibles d’entraîner les graines/fruits d’une tache à l’autre ou vers d’autres taches non connectées. Le suivi floristique des taches confirme d’ailleurs ces constatations à propos du vent puisque sur les douze ans de suivi les taches connectées ont gagné 15% de plus d’espèces dispersées par le vent que les taches ailées ou rectangulaires.
En pratique

Pulsatille rouge des pelouses granitiques rases 
Fruits plumeux transportés à longue distance … en absence d’obstacle !
Cette étude très théorique en apparence n’en a pas moins des prolongements pratiques en matière de conservation et de gestion des habitats ouverts fragmentés menacés par la reforestation massive : la création ou le maintien de corridors linéaires connectant le plus possible les taches entre elles semble bien déterminant pour la survie des espèces liées à ces milieux. Cette nécessité devient encore plus forte si l’on ajoute le problème de la crise climatique en cours : les plantes (et les animaux) doivent avoir la possibilité de circuler pour avoir des chances de trouver de nouveaux sites climatiquement convenables. L’étude apporte une préconisation pratique supplémentaire : si on créé un corridor, il faut aligner son axe sur la direction des vents dominants ce qui va optimiser les trois effets mis en évidence et notamment favoriser les points chauds d’éjection et la dispersion à longue distance, capitale pour la survie des espèces. De la même manière, lors d’opérations de restauration de tels milieux par réouverture du milieu forestier, il convient d’orienter les parcelles créées dans la direction des vents dominants.

On aurait pu croire a priori que le vent en tant qu’agent de dispersion n’était pas impacté par les transformations des paysages mais cet exemple montre qu’à l’échelle des petites taches isolées les effets de lisière changent tout et agissent directement sur la dispersion des plantes anémochores liées aux milieux ouverts.

Aster amelle : espèce patrimoniale des coteaux secs en fort déclin avec l’enfrichement des pelouses et les plantations de résineux 
Fruits plumeux de l’aster amelle
Bibliographie
How fragmentation and corridors affect wind dynamics and seed dispersal in open habitats.Ellen I. Damschen et al. 3484–3489. PNAS. 2014 ; vol. 111 ; no. 9.
