Le mélampyre des champs ou blé de vache
Melampyrum arvense

10/06/2023 Le mélampyre des champs fait partie de ces plantes impossibles à rater et très faciles à identifier tant elles s’imposent dans le paysage par des traits uniques dont les vives couleurs des inflorescences très voyantes. Dommage qu’il soit en forte régression dans les campagnes du fait de l’intensification agricole : il constituait un beau marqueur de paysages agricoles équilibrés en colonisant les bords des champs des céréales. Cette espèce se démarque dans trois domaines : la pollinisation de ses fleurs, la dispersion de ses graines surprenantes et son mode de vie d’hémiparasite.

Rougeole

Le mélampyre des champs ne manque pas de surnoms populaires originaux dont une partie résulte de l’aspect très original de ses floraisons de mai à juillet (jusqu’en août plus rarement) : rougeole, rougeote, rougette, herbe rouge, saponaire rouge ou encore queue de renard. Ils font tous allusion à la coloration rouge carmin profond de ses longs épis denses, dressés et presque cylindriques.



En s’approchant, on découvre que cette teinte dominante ne provient pas en fait des fleurs elles-mêmes mais des sortes de feuilles qui les accompagnent : des bractées disposées sur deux rangs. Recourbées en pointe dressée, elles ont des bords découpés profondément en lanières pointues et sous-tendent les fleurs. Elles sont tellement serrées qu’elles cachent l’axe de l’épi (le rachis).
Les fleurs frappent par leur aspect bariolé et leur forme de « gueule de loup » fermée (fleur irrégulière ou zygomorphe). Longue de 2 à 2,5cm, la corolle dépasse nettement du calice poilu peu visible avec quatre dents presque égales. Elle se compose de deux parties : un tube allongé de presque deux centimètres de long émerge du calice mais se coude fortement à sa base ce qui le rend presque horizontal ; puis, le tube s’élargit en « gueule » et se ferme par deux lèvres en forme de bourrelets : l’inférieure est pressée contre la supérieure en forme de capuchon aplati latéralement. D’un rose magenta foncé, finement veloutées, les corolles arborent une large bande jaune en travers à mi-parcours.
L’ensemble bractées/fleurs forme comme une belle pagode bariolée avec des fleurs regardant dans toutes les directions ; comme de plus, ce mélampyre forme presque toujours des colonies denses de nombreux individus, l’effet coloré est saisissant.


Pour le reste, le mélampyre est une plante dressée à tige principale simple portant des étages successifs de rameaux écartés redressés avec des feuilles opposées dentées découpées en lanières à leur base (comme les bractées au-dessus).
Fleur à bourdons
Sous le capuchon de la corolle (galea) se logent les anthères des deux paires d’étamines opposées dont la base se prolonge par une pointe. L’ovaire logé au fond du calice porte un style terminé par un stigmate saillant. Sous cet ovaire, au plus profond du tube de la corolle, se trouve une glande à nectar (nectaire) qui produit un abondant nectar (plante très mellifère). Pour y accéder depuis l’entrée fermée, un insecte visiteur doit donc disposer d’une langue-trompe d’au moins 2cm de long ! Chaque fleur dure en moyenne sept jours.
Les principaux visiteurs sont des petits bourdons de diverses espèces : ils sont pratiquement les seuls capables « d’ouvrir la fleur ». Il faut en effet appuyer fortement sur la lèvre inférieure en forme de plateforme pour entrouvrir la « gueule de loup » et y introduire sa tête avec la langue-trompe déployée pour atteindre le nectar convoité. Seules les espèces à langue longue peuvent ainsi accéder au nectar de manière légitime.

Bourdon avec sa langue déployée 
Bourdon butinant par l’entrée de la fleur 
Les bourdons à langue longue ou moyenne récupèrent du pollen sur la tête et le thorax car ils appuient leur tête dans la chambre pollinique du capuchon ; après plusieurs visites, ils se toilettent et amassent ce pollen sur les brosses à pollen de leurs pattes postérieures. Pendant la phase de récolte du nectar, une partie de ce pollen pourra atteindre le stigmate d’une fleur visitée. Ils visitent même les fleurs encore en bouton : mais comme le stigmate est déjà réceptif, ils peuvent ainsi assurer la pollinisation s’ils portent du pollen d’autres fleurs.

Corolle découpée : anthères sous le capuchon de la lèvre supérieure 
Les étamines se retrouvent unies à la pointe des anthères
Ces fleurs sont autofertiles : théoriquement, leur pollen peut germer sur leur propre stigmate et assurer la fécondation. Mais si on encage les fleurs en les privant de visiteurs, elles ne produisent pas de graines car le stigmate ne réussit pas à recevoir du pollen. Donc, le succès reproductif des populations de mélampyre des prés dépend étroitement de la présence de pollinisateurs efficaces ; si la végétation environnante devient trop haute par exemple, elle limite l’accès aux fleurs et la fructification s’effondre. Si on pollinise manuellement (avec un pinceau) de manière croisée des fleurs, cela augmente la fructification et le nombre de graines par fruit augmente ce qui confirme bien l’importance des pollinisateurs.
Efficacité

Mais, les espèces de bourdons à langue courte (comme le bourdon terrestre ou des prés) arrivent aussi à collecter ce nectar de manière « illégitime » du point de vue de la fleur. Ils trichent de deux manières. Pour récolter le pollen, ils s’accrochent sous la lèvre et mordent les bords pour la dégager : en faisant vibrer leurs ailes, ils font tomber le pollen des anthères découvertes qui tombe cette fois sur leur ventre (ils sont en position inversée) ; ils peuvent éventuellement déposer un peu de pollen sur le stigmate mais avec moins d’efficacité. Pour récolter le nectar, avec leur langue courte mais robuste, ils percent un trou à la base du tube au niveau du coude et en introduisant leur langue accèdent ainsi au nectar. Ce faisant, ils n’assurent pas la pollinisation puisque leur corps éventuellement porteur de pollen n’entre pas en contact avec le stigmate : on parle de tricherie !
D’autres bourdons à langue courte ou des abeilles domestiques utilisent ensuite les trous percés par les tricheurs et se comportent donc en tricheurs secondaires. Dans une population étudiée, presque 80% des fleurs avaient de tels trous et 20% d’entre elles avaient leur pistil détruit par ces morsures du tube. 44% des fleurs visitées par des bourdons à langue longue sont pollinisées contre seulement 25% pour ceux à langue courte.

La densité des colonies influe largement sur les taux de visite mais de manière contre-intuitive : les plantes en faible densité adoptent un port plus ramifié (voir ci-dessous) et portent de ce fait plus de fleurs par plante ; les fleurs des rameaux latéraux produisent plus de graines par fleurs. Les fleurs sont visitées en moyenne 6 à 7 fois durant leur semaine de floraison mais, en conditions favorables, ce taux peut passer à 40 fois !
Fleur à fourmis
Les fleurs fécondées se transforment en fruits secs : des capsules glabres, d’environ 1cm de long, cachées dans les calices. Chaque capsule se compose de deux loges renfermant chacune une seule graine. Grosse (6mm de long sur 2 à 3 de large) et lisse, elle présente à maturité une teinte brun jaune et rappelle nettement un grain de blé (voir le paragraphe suivant).

Capsule verte (calice déchiré) 
Graines extraites 
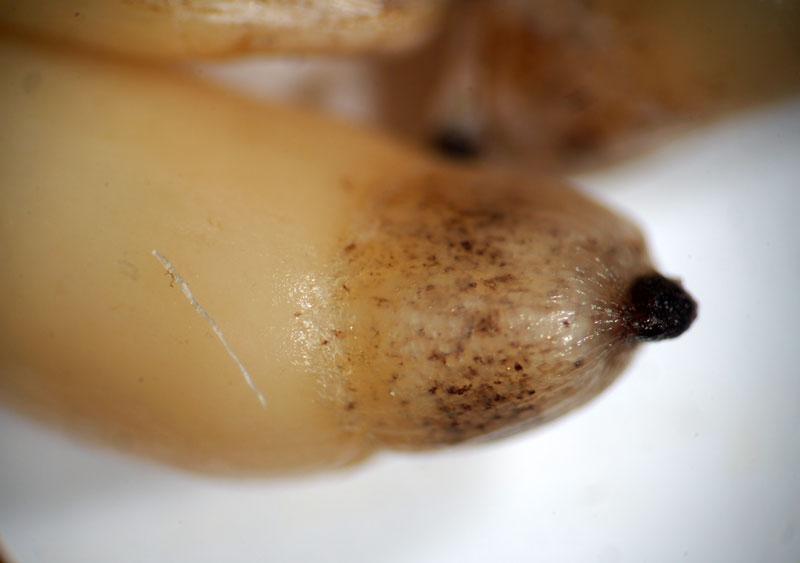
Elaïosome au sommet d’une graine
La graine porte à une extrémité un appendice épais un peu charnu : un élaïosome, riche en lipides et très recherché des fourmis. Celles-ci récoltent les graines tombées au sol après ouverture des capsules sèches en les saisissant par cet élaïosome : arrivées à leur fourmilière, elles le détachent pour l’emporter comme nourriture pour leur couvain ; la graine détachée est rejetée à l’entrée. Il s’agit donc d’un mode de dispersion à courte distance (quelques mètres au plus le plus souvent) qualifié de myrmécochorie (myrmeco : fourmi).

Plantules rougeâtres 
Globalement, même avec cette aide animale, le mélampyre des champs souffre d’un certain manque de dispersion de ses graines : elles tendent à s’accumuler au pied des plantes mères. Ainsi, au printemps, les plantules se développent souvent en paquets denses. On pense que néanmoins ceci favorise le développement de suçoirs (haustorias) au niveau des racines parasites par effet de compétition ce qui améliorerait finalement la prise d’eau et de nutriments aux dépens des plantes hôtes.

Points noirs sous les bractées : nectaires extrafloraux 
Fourmi en train de visiter un nectaire
L’interaction positive avec les fourmis ne se limite d’ailleurs pas à la seule dispersion des graines. Les bractées des inflorescences (voir ci-dessus) sont marquées de forts points noirs qui sont en fait des nectaires, des glandes sécrétrices de nectar ; on parle de nectaires extra-floraux. Les fourmis attirées par cette production viennent lécher ces nectaires et protègent sans doute indirectement la plante des attaques d’herbivores. Au passage, elles doivent peut-être aussi récolter les graines quand les premières capsules sont mûres alors que le sommet de l’épi floral est toujours en fleur ?
Blé de vache
La ressemblance des graines avec des grains de blé a de quoi intriguer quand on sait qu’autrefois le mélampyre des prés abondait dans les moissons de céréales (voir ci-dessous), son habitat secondaire. En plus, la plante fructifie et mûrit en début d’été soit en même temps que la moisson. Ceci lui a valu divers noms populaires : blé ou froment de vache (repris en anglais : cow-wheat), blé noir, millet sauvage, …
On pourrait penser qu’il s’agit là d’un exemple de mimétisme végétal où une plante en imite une autre et en tire profit pour sa dispersion : plusieurs adventices des cultures ou messicoles dont la nielle des blés ou la folle-avoine procèdent ainsi. Mais, ce trait de ressemblance se retrouve chez les autres mélampyres ne vivant pas dans ce milieu dont le mélampyre des prés (nom trompeur : espèce surtout forestière !). D’ailleurs le nom de genre mélampyre (Melampyrum) vient de deux racines : melas, noir (voir la mélanine) et puros, blé. Il se peut par contre que le mélampyre des champs ait bénéficié la présence de ce trait pour coloniser efficacement de manière secondaire les cultures, de manière fortuite : l’Homme aurait alors contribué à le propager avec les semences.

Ses graines se retrouvaient donc facilement dans les récoltes de céréales à une époque où n’existait pas de dispositif de tri des graines. Elles donnaient à la farine et au pain contaminé une teinte rougeâtre et rendaient leur consommation dangereuse : elles renferment une substance toxique dangereuse, l’aucubine, un iridoïde qui existe aussi par exemple chez le ginkgo.
Dans le sud de l’Angleterre où le mélampyre des champs était très commun dans les moissons, on le surnommait de ce fait « l’herbe de la pauvreté » : il dépréciait considérablement la valeur marchande du blé contaminé. Sur l’île de Wight, en 1823, le Dr Broomfield étudia soigneusement son histoire :
« La tradition locale voulait que les graines aient été importées de « régions étrangères » (certains disaient d’Espagne, d’autres de Jersey, d’autres, qui avaient plus probablement raison, du Norfolk). Une coutume locale consistait au moment de la récolte à arracher cette herbe avec grand soin et de l’emporter dans des sacs, de la brûler, et de ramasser les moindres graines qu’on pouvait voir au sol. Le pain, fait à partir du blé des fermes autour de Undercliff (sud de l’île) était ainsi moins foncé et « chaud » qu’il ne l’était auparavant car cette plante diminuait en nombre. »
Sur le déclin
Le mélampyre des champs ne se trouve que sur des sols calcaires crayeux ou marneux, secs (pouvant être humides en hiver mais se desséchant au printemps) et en pleine lumière ; il a besoin de chaleur et dans la moitié nord du pays se cantonne sur les pentes bien exposées.

Colonie en lisière (ourlet) 
Talus en bordure d’un bois
Son habitat primaire originel se situe sur les lisières chaudes des boisements : les ourlets herbacés ; là, il côtoie le plus souvent d’autres espèces thermophiles (aimant la chaleur) comme lui et apportant chacune une note colorée vive à la hauteur de celle du mélampyre : coronille variée, géranium sanguin, trèfle rouge, vesce à feuilles ténues, …


Pelouse basse et maigre 
Pelouse en voie d’enfrichement 
Pelouse pâturée par des chevaux (voir la clôture)
Secondairement, il a donc colonisé (voir ci-dessus) les moissons de céréales sur sol calcaire tout en restant essentiellement sur les bordures. Au moins trois techniques « modernes » de gestion des cutures de céréales l’en ont quasiment éliminé : les herbicides évidemment, le brûlage des chaumes et le tri des graines (voir ci-dessus). Il s’est cependant réfugié dans les friches calcaires après abandon des cultures où il ne persiste que quelques années tant que la végétation herbacée n’est pas trop haute ou sur les accotements et les talus des bords de routes … non traités ni fauchés ou broyés de manière récurrente ! Sa présence au moins sur ces derniers dans les plaines agricoles pourrait facilement être facilitée en allégeant les modes de gestion de ces tiers milieux : le mélampyre des blés apporte dans le paysage agricole une superbe note colorée et offre ses fleurs aux pollinisateurs dont les bourdons (voir ci-dessus).
Actuellement, cette espèce se retrouve donc très dispersée dans toute la France jusqu’à 1700m, loin de son statut ancien d’espèce très commune ! En Grande-Bretagne (voir ci-dessus), de très commun autrefois, il se retrouve désormais localisé dans moins d’une dizaine de sites du sud de l’Angleterre.
Hémiparasite

Orobanche du panicaut 
Lathrée clandestine
Le mélampyre des champs appartient à la famille des Orobanchacées qui ne regroupe que des espèces parasites sur les racines de plantes hôtes : soit entièrement (holoparasites) et alors dépourvues de chlorophylle comme orobanches et lathrées, soit en partie (conservant la fonction chlorophyllienne : hémiparasites) comme les rhinanthes, les euphraises, les odontites, … et les mélampyres.

Odontite rouge 
Rhinanthe 
Euphraise
Deux signes permettent de détecter ce mode de vie qui passe inaperçu autrement. Si on arrache un pied de mélampyre, il sèche très rapidement et devient tout noir notamment dans les planches d’herbier. Ce « symptôme » indique sa dépendance envers son hôte. D’autre part, sur le pied arraché, on remarque que l’appareil racinaire est très réduit au regard de la taille de la plante.

Plante sèche noircie 
Racine très rudimentaire
Les plantules issues de la germination des graines produisent une petite racine qui se ramifie jusqu’à rencontrer celle d’une autre plante : elle établit alors des connections en émettant des suçoirs ou haustorias, structures communes à toutes les plantes parasites (voir les cuscutes par exemple). Si la plantule ne trouve pas d’hôte dans les semaines suivant sa germination, elle meurt.

Le mélampyre des champs (comme les autres mélampyres) se montre très peu sélectif dans son choix de plantes hôtes : il peut se développer aux dépens de plantes de familles très diverses, y compris en culture sur des plantes très toxiques (belladone, ricin, … !). Un individu peut parasiter plusieurs espèces en même temps même si toutes ne lui profitent pas de la même manière. Il croît plus vite sur des légumineuses (riches en azote) ou des graminées. On a démontré que les excrétions racinaires de ces dernières stimulaient les suçoirs des mélampyres : ceci explique sans doute la colonisation historique des champs de céréales (voir ci-dessus).
Le mélampyre prélève de l’eau, des nutriments et des composés secondaires des plantes hôtes dont d’éventuelles substances toxiques. On a testé l’appétence du mélampyre des champs selon les plantes parasitées pour deux invertébrés herbivores : la chenille de la noctuelle du cotonnier (bioagresseur des cultures) et une petite limace. Les mélampyres testés ont été cultivés sur 19 espèces hôtes de 11 familles différentes connues pour contenir un large spectre de composés secondaires toxiques. Les deux herbivores testés réagissent différemment mais se montrent chacun capable de discriminer la qualité nutritive du mélampyre selon ses hôtes parasités. Par contre, l’appétence de l’espèce végétale hôte du mélampyre n’intervient pas dans la préférence ou pas des herbivores. Autrement dit, le mélampyre n’endosse que partiellement les qualités intrinsèques de ses hôtes. Ceci va à l’encontre de croyances anciennes selon lesquelles le parasite « devenait » l’hôte du fait qu’il s’en nourrissait.
Bibliographie
Flora Britannica. R. Mabey. Ed. Chatto and Windus.1996
Pollination ecology and seed-set in the rare annual species Melampyrum arvense L. (Scrophulariaceae). M.M.Kwak Acta Bot. Neerl.37(2), 1988,p. 153-163
Host-parasite relations in the root hemiparasite Melampyrum arvense. D. Matthies. Flora (1995) 190. 383-394
Is palatability of a root-hemiparasitic plant influenced by its host species? M. Schädler et al. Oecologia (2005) 146: 227–233
Interactions between the Root Hemiparasite Melampyrum arvense and Mixtures of Host Plants: Heterotrophic Benefit and Parasite-Mediated Competition D. Matthies Oikos Vol. 75, No. 1 (Feb., 1996), pp. 118-124
