Les nids des oiseaux (2) : des signaux sexuels
Nous avons vu dans une première chronique sur les nids l’importance de la pression de sélection naturelle sur la conception des nids en lien avec les risques liés à la prédation. Les nids, construits sciemment par les oiseaux, représentent en quelque sorte “un prolongement d’eux-mêmes », une expression extérieure qui leur permet dans ce cas de se reproduire. Les scientifiques parlent à ce propos de phénotype étendu, i.e. que le nid « fait partie de l’oiseau en quelque sorte ».
Via la conception de son nid, le bâtisseur/la bâtisseuse (ou les deux selon les espèces) peut aussi envoyer des signaux lisibles sur sa qualité de reproducteur(trice) comme il/elle peut l’exprimer aussi à travers divers signaux corporels (taches colorées, ornements, …) ou comportementaux (chant, parade, …) : voir l’exemple du chant de la huppe. Les traits des nids faisant office de signaux sexuels se trouvent soumis de leur côté à la pression de la sélection sexuelle qui vient se surajouter à la sélection naturelle anti-prédation évoquée dans la première chronique. Ils seront l’objet de cette nouvelle chronique.
Coûts de construction
Dans la théorie sur la sélection sexuelle, il est postulé qu’un trait sélectionné doit être un facteur de risque relatif pour celui/celle qui le porte : sa capacité à surmonter ce handicap. Donc pour jouer un rôle dans la sélection sexuelle, la conception et la construction des nids doivent présenter pour celui/celle qui le fait un certain coût (risques, énergie dépensée, …). On a longtemps minimisé ces coûts en mettant plutôt l’accent sur la ponte et l’élevage des jeunes qui suivent. Les dernières décennies de recherche ornithologique ont au contraire démontré qu’il n’en était rien comme l’indiquent d’ailleurs, par exemple, la fréquence des scènes de « vol de matériaux » dans les nids de congénères ou les espèces qui nichent dans d’anciens nids d’autres espèces.

Hirondelle rustique : des centaines de boulettes de boue récoltées et transportées … 
… pour édifier une coupe qui peine à contenir les grands jeunes !
On a démontré chez les hirondelles rustiques que plus elles déployaient d’efforts pour bâtir leurs nids faits de boulettes de boues agglomérées, plus leur système immunitaire était réactif (évalué à partir de prises de sang).
Expérimentalement, on a testé les coûts associés à la construction du nid chez le gobe-mouches noir, passereau cavernicole forestier. On enlève les nids et œufs de couples expérimentaux en cours de nidification ce qui les amène à en construire un second pour effectuer une ponte de remplacement. Alors, les femelles construisent des nids plus petits que les premiers initialement construits. Inversement, si on abaisse expérimentalement les coûts de construction, les femelles concernées passent plus de temps à couver les œufs et ravitaillent ensuite à un rythme plus élevé leur progéniture. Ainsi, les jeunes issus de telles nichées, mesurés à l’envol, ont des pattes plus grandes ce qui laisse penser qu’ils ont mieux grandi. De même chez les mésanges bleues, si on enlève expérimentalement le nid et les œufs alors que les femelles couvent depuis cinq jours, les femelles ainsi « maltraitées » ( !), reconstruisent un nouveau nid plus petit et pondent moins d’œufs dans la ponte de remplacement.

Mâle de gobe-mouches noir (Cliché ViruPersonen ; C.C.4.0.) 
Gobe-mouches noir installé dans un nichoir artificiel ( Cliché RobStoeltje ; C.C.2.0.)
Donc, assez clairement, oui construire un nid « pompe » suffisamment d’énergie pour influer sur le succès reproductif ultérieur et donc les comportements associés et l’architecture du nid constituent bien des signaux fiables de l’état physique et physiologique de ou des individus bâtisseurs.
Voyons donc ce qui se passe selon qui est le sexe bâtisseur.
Nids masculins
Divers exemples confirment que, chez les espèces où le mâle est le bâtisseur, il existe bien un lien positif entre la taille et/ou la qualité de construction du nid et certains aspects de sa constitution physique et physiologique.
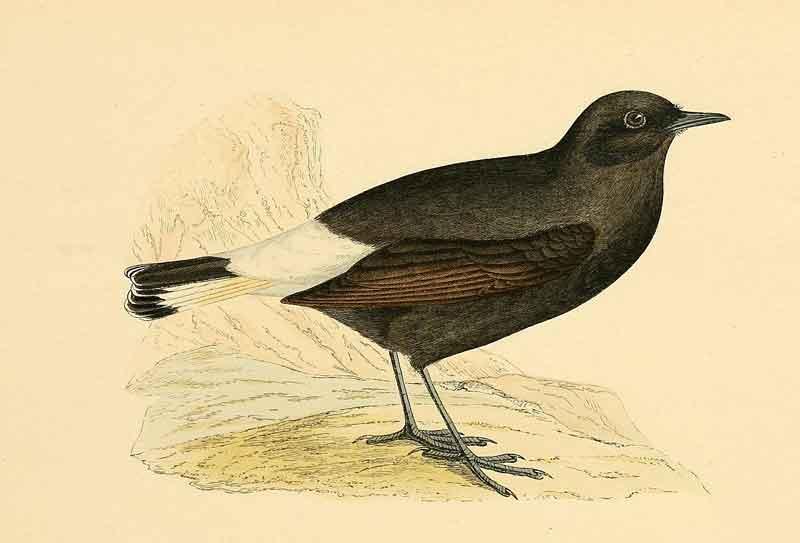
Premier exemple un peu édifiant (jeu de mots !) : le traquet rieur, sédentaire en Afrique du Nord et Espagne avec quelques couples sur la côte des Pyrénées orientales. Ce passereau niche dans des sites rocailleux arides ; le mâle installe le nid (coupe d’herbes et de radicelles) dans un trou de rocher ou de mur.

Mais, il construit en plus, devant l’entrée, une barrière de cailloux qu’il récolte autour ; il les accumule au fil des années où il occupe le site. Cette barrière constitue donc un signal visuel qui s’impose dans le paysage. Il transporte des cailloux de 3 à 10 grammes en moyenne mais parfois jusqu’à … 28 grammes (soit les 2/3 de son propre poids) ! On estime qu’au cours d’une saison, un mâle transporte au total … 2 kg de pierres ! Les mâles avec des surfaces d’ailes plus grandes transportent plus de pierres que ceux à ailes plus petites. Expérimentalement, une année donnée, alors que le mâle a commencé à entasser de nouvelles pierres, on enlève les pierres des années précédentes : contre toute attente le mâle n’en rajoute pas plus de nouvelles pour compenser ; ceci signifie que les femelles choisissent les mâles sur la base du nombre de pierres nouvelles apportées cette année-là. Inversement, si on ajoute de nouvelles pierres à la barrière en construction, le mâle en transporte moins ! Enfin, on peut manipuler le nombre de pierres neuves par nid : les femelles appariées avec les mâles ayant au final le plus de pierres nouvelles pondent plus tôt en saison de reproduction et, de ce fait, ont un meilleur succès reproductif.

Etourneau unicolore (Cliché J. Prego ; C.C. 2.0.) 
Etourneau unicolore nichant dans une cavité d’arbre (Cliché L. Worthington ; C.C. 2.0.)
Second exemple bien particulier : l’étourneau unicolore, espèce proche de l’étourneau sansonnet mais méditerranéenne (nicheur en Corse et Pyrénées-Orientales). Chez les étourneaux, c’est le mâle qui bâtit le nid dans une cavité ; la femelle se contente d’y ajouter quelques plumes à la fin. Les mâles incorporent dans la coupe du nid des rameaux de végétaux verts frais. L’essentiel de cet ajout se fait dans les dix jours avant le début de la ponte. Si on ajoute ou enlève expérimentalement ces végétaux verts frais, cela ne change rien dans la charge en parasites des jeunes au nid ni le poids des jeunes à l’envol : ceci indique qu’ils n’auraient donc pas de fonction antiparasitaire comme on pourrait le penser. Par contre, les mâles avec plus de matériaux verts attirent avec plus de succès des femelles lors de la formation des couples ; les femelles réagissent alors en apportant plus de plumes. Si on ajoute des matériaux verts, les femelles pondent plus d’œufs et les nichées comportent plus de mâles que de femelles à l’éclosion ! Tout ceci confirme la puissante valeur de « signal sexuel » de ces matériaux.

Dans diverses espèces, les mâles bâtissent plusieurs nids en début de saison et les femelles choisissent leur partenaire sur la base du nombre de nids proposés. Tel est le cas bien connu du minuscule troglodyte mignon : la femelle, une fois son choix de mâle effectué, sélectionne un des nids et en complète le revêtement intérieur.
Nids féminins
On connaît assez peu d’espèces où seule la femelle bâtit le nid (voir nids biparentaux) si bien que ce cas a été assez peu étudié.
Les femelles d’étourneau unicolore (voir ci-dessus) ne placent pas les plumes au fond du nid au hasard : les plumes avec une forte réflectance aux UV sur leur face externe sont presque systématiquement installées cette face vers le haut. Par contre, si elles récoltent des plumes de geai pour lesquelles la réflectance maximale est sur l’autre face, elles les tournent de côté ; les plumes de pie bleue (espèce espagnole) qui réfléchissent autant des deux côtés sont placées indifféremment ! Donc, les femelles sélectionnent la position qui rend ces plumes les plus visibles aux yeux du partenaire.

Diversité des matériaux du nid des mésanges bleues (Cliché R. Waller ; C.C. 4.0.) 
(Cliché nottsexminer ; C.C. 2.0) 
(Cliché Aruminska ; C.C. 2.0.)
Les mésanges bleues femelles les moins infectées par un parasite sanguin répandu bâtissent des nids plus lourds que les femelles plus infectées. Certains mâles apportent aussi quelques plumes en fin de construction (situation inverse de l’étourneau) ; or, ces mâles ont des pattes plus longues et participent plus activement au nourrissage des jeunes : les femelles appariées avec ceux-ci réagissent à l’apport de plumes en participant moins au nourrissage. Les jeunes élevés par de tels couples (mâle plumassier !) sont en meilleure forme physique au moment de l’envol. Si on ajoute des plumes au nid d’une femelle qui en a déjà reçu, imitant ainsi un cas d’apport par un mâle extra-couple, les mâles concernés réagissent en abaissant leur taux de nourrissage !

Les femelles de mésanges bleues pratiquent aussi l’apport de matériaux verts au nid (voir l’étourneau ci-dessus) et cette fois ils peuvent avoir des effets antiparasitaires (voir l’exemple des mésanges bleues de Corse et l’épisode 3). Dans une étude expérimentale, on a fait varier la quantité de matériaux verts déposés dans le nid et on observe la réaction du mâle. Sur les nids dont a diminué la quantité de matériaux, les mâles adoptent moins de comportements à risques (par exemple vis-à-vis des prédateurs). Les mâles qui prennent plus de risques pour les nids enrichis en matériaux verts voient le succès reproductif de leur couple amélioré. Autrement dit, en plaçant plus de tels matériaux, les femelles augmentent leur succès reproductif via un investissement accru du mâle.
Nids biparentaux
Avec les nids masculins, ce sont les cas plus répandus chez les oiseaux mais pour autant ils ont été moins étudiés car plus complexes.

Chez la mésange huppée, seuls les mâles en bonne condition physique participent à la construction du nid ce qui raccourcit alors le temps de mise en place et permet à la femelle de pondre plus tôt (5 jours en moyenne). Les jeunes émergent donc 5 jours plus tôt ce qui leur donne plus de chances de succès à l’envol. La participation du mâle améliore donc le succès reproductif du couple.

Les mâles d’hirondelles rustiques avec de longs filets de la queue, signe associé à une meilleure qualité physique, contribuent moins à la construction du nid. L’énergie dépensée par la femelle dans le bâtissage du nid reste la même quel que soit le type de mâle (collaborant ou pas) ; mais les nids où le mâle ne collabore ont des parois plus minces tout en étant plus grands : ils peuvent ainsi héberger des nichées plus importantes.
Pour autant, ces signaux peuvent parfois être « détournés » et en pas indiquer de manière honnête l’état réel du bâtisseur. Ainsi chez le milan noir, on sait que l’ajout d’objets hétéroclites d’origine humaine (dont des sacs plastiques blancs : voir la chronique) signale, de manière honnête, la capacité du couple bâtisseur à défendre le nid contre les intrusions. Mais des milans non nicheurs qui n’ont pas réussi à s’installer sur des territoires offrant des sites de nids tentent de manière assez violente de s’en emparer ; la présence de nombreux déchets dans le nid du territoire occupé les avertit de la haute qualité défensive du couple déjà installé. Même les couples de « basse qualité » défensive ne trichent pas et mettent peu de déchets dans leur nid : ils pourraient très bien le faire après tout mais on pense qu’ils craignent ainsi de s’exposer à des attaques virulentes de la part d’intrus ! La pression est donc grande pour ne pas tricher et maintenir « l’honnêteté » du signal.

Nids de pie bavarde 
Chez les pies bavardes, les deux parents construisent le grand nid avec son toit en dôme typique. On a observé que les femelles ajustent leur investissement reproductif au degré de participation du mâle à la construction. Les couples de haute qualité bâtissent de ce fait des nids plus grands. Si on enlève la première nichée éclose et le nid pour de tels couples, ils se montrent plus aptes à en rebâtir un nouveau ; de plus, les femelles pondent plus d’œufs dans les nids construits plus vite (donc avec un mâle très collaboratif).

Jeune coucou-geai en compagnie de son parent « adoptif » (une corneille mantelée) (Cliché R. Silberman ; C.C. 4.0) 
L’oeuf du coucou-geai (à gauche) ressemble fortement à ceux de la pie bavarde (les deux à droite) (Cliché Museum Wisbaden ; C.C. 3.0)
Il se trouve que, dans le Midi, les nids de pies (et d’autres corvidés comme les corneilles) sont souvent parasités par une espèce de coucou, le coucou-geai : il dépose ses œufs dans les pontes de pie et celles-ci les couvent et élèvent les jeunes. Les coucou-geais choisissent de préférence les nids plus grands a priori tenus par des couples de bonne qualité. En retour, dans les secteurs où sévit ce parasite de nichée, les pies tendent à bâtir des nids plus petits : une autre pression de sélection !
Le cas des moineaux domestiques complète cette idée que différentes formes de sélection s’exercent en même temps. Les mâles moineaux ajoutent des plumes au nid mis, quand ils le font, ils crient beaucoup pour attirer l’attention des femelles. Si on enlève les plumes ajoutées, les mâles en ramènent rapidement de nouvelles. Le volume de plumes apportées au nid est lié à la taille de la ponte et au rythme de nourrissage des poussins par la femelle. Les plumes sont ajoutées pendant l’incubation et en début de période d’élevage des jeunes, soit au moment où elles peuvent apporter un certain avantage d’isolant thermique (effet couette en duvet !). De ce fait, ce comportement des mâles subit à la fois des pressions de sélection naturelle (meilleur succès des jeunes) et de sélection sexuelle (voir ci-dessous) !

Mâle de moineau domestique avec des plumes blanches fièrement arborées (cliché P Jeganathan ; C.C. 4.0) 
Mâle « qui veut » frapper l’attention des femelles » avec un vieux kleenex ! (cliché Lee ; C.C. 2.0)
Ces exemples indiquent clairement que les nids représentent bien un indicateur fiable de la qualité du ou des parents bâtisseurs. Les nids contiennent les œufs et/ou la nichée (selon que les jeunes restent au nid ou pas) : ainsi, ils sont soumis aussi bien aux pressions de la sélection naturelle qu’à celles de la sélection sexuelle. La première favorise les nids plus petits (anti-prédation) alors que la seconde favorise la « mégalomanie » pour attirer l’attention et signaler sa bonne qualité ! Ainsi, l’évolution des nids et de leur construction se fait sans cesse dans le compromis.
A noter que les structures très élaborées et hautement artistiques des oiseaux-berceaux, qui font penser à des nids, ne servent qu’à attirer les femelles pour s’accoupler : dans ce cas, ces « faux nids » sont uniquement des signaux sexuels. Les femelles accouplées vont ensuite, seules, bâtir un nid ailleurs, pondre et élever les jeunes.
Rendez-vous vers la troisième chronique consacrée aux nids comme variables d’ajustement à l’environnement.
Retrouvez toutes les chroniques sur les nids d’oiseaux (dont ces quatre épisodes) ici
Bibliographie
1)Bird nests and construction behaviour. Review. M. Hansell. Ed. Cambridge U.P. 2000
2) The design and function of birds’ nests Mark C. Mainwaring et al. Ecology and Evolution 2014; 20(4): 3909– 3928
3)Nest predation research: recent findings and future perspectives. Review J. D. Ibanez-Alamo et al. J Ornithol (2015) 156 (Suppl 1):S247–S262
