L’Érythrone dent-de-chien
Erythronium dens-canis


23/03/2024 Érythrone dent-de-chien : un nom étrange qui n’inspire guère a priori. Et pourtant, une rencontre avec des érythrones relève de l’enchantement tant leurs fleurs affichent une beauté insolente. Mais, les chances de rencontre avec cette fée envoûtante restent limitées car l’espèce a une aire de répartition limitée et sa fenêtre de floraison est très brève au printemps. Ses fleurs captent irrésistiblement l’attention de l’observateur subjugué. Mais, si on détourne juste un peu son regard pour l’observer dans sa globalité et la suivre dans son bref cycle de vie, on découvre une série de détails inattendus et passionnants ; comme la belle a fait l’objet de nombreuses études scientifiques, celles-ci nous apportent d’autres révélations sur de la vie des érythrones dont une sidérante livrée à la toute fin de cette chronique, exceptionnellement longue !
Fleur sublime
Chaque pied individuel porte au sommet d’une hampe florale (tige nue) dressée une seule fleur : mais quelle fleur ! Longue de 2 à 3,5cm, elle est penchée en avant : six tépales rose pourpre, recourbés vers l’extérieur de la plus belle des façons. On parle de tépales car sépales et pétales ont la même consistance et se retrouvent associés dans un même cercle (périgone), un caractère que l’on retrouve chez les autres membres de sa famille, les Liliacées, comme les lis, les tulipes, les gagées ou les fritillaires.

Hampe florale avec sa fleur 
Fleur penchée : noter le stigmate blanc qui dépasse les étamines 
Fleur dont on a enlevé un tépale 
Anthères colorées des étamines saillantes
Chaque tépale, au niveau de la courbure, porte une large tache blanc et jaune foncé. Les bases droites des six tépales très rapprochés forment un tube allongé au fond duquel est secrété un abondant nectar.
Du centre de la fleur surgissent six étamines fortement saillantes à l’extérieur et le pistil blanc. Chaque étamine porte une longue anthère de teinte violet clair qui vire au bleu foncé à maturité et libère un pollen très coloré. Le pistil qui commence avec l’ovaire caché au fond du tube floral se compose d’un style allongé terminé par un gros stigmate à trois branches à peine écartées.
On notera que ce stigmate se trouve sensiblement en avant des anthères (plus ou moins !) : on appelle herkogamie (herkos, clôture) ce décalage spatial entre étamines et stigmate,ce qui limite les probabilités d’autofécondation en recevant du pollen de ses propres étamines.
Pollinisation
Des fleurs aussi voyantes ne manquent pas d’attirer des insectes pollinisateurs … pour peu que les conditions météorologiques leur soient favorables. En effet, l’érythrone fleurit tôt : dès mars à basse altitude et jusqu’en juin pour les stations alpines (2200m) ; souvent, le déclencheur est la fonte des neiges sur les sites enneigés en hiver.

Bourdon en cours de visite : noter le style tordu tant l’insecte est actif ! 
Il tourne sui lui-même et sonde avidement le tube floral
Dans le nord-ouest de l’Espagne, les visiteurs principaux observés sont les grosses reines des bourdons terrestres sorties d’hibernation et divers andrènes, des abeilles solitaires ; ailleurs, ce sont les abeilles domestiques qui dominent (45% des visites) suivies d’autres hyménoptères (33%) et des mouches. Les bourdons semblent tout particulièrement apprécier ces fleurs et passent un temps anormalement long sur chaque fleur, en position renversée, et tournant dans tous les sens frénétiquement pour trouver des accès au nectar avec leur longue langue passée entre les tépales.

Les taches jaunes observées sur les tépales (voir ci-dessus) servent de guides à nectar : elles orientent les insectes visiteurs vers l’entrée de la fleur. Or, on sait que les bourdons ont une vision plus étendue que la nôtre en termes car ils perçoivent les UV ; on peut donc supposer que, vues sous un éclairage UV, ces taches doivent bien plus ressortir et devenir des sémaphores contrastés.
Des expériences in situ montrent que 74% des fleurs en libre accès sont effectivement pollinisées et fructifient ensuite : la production de fruits reste donc forte en situation naturelle. Par contre, si on empêche l’accès des visiteurs, seules 21% sont pollinisées ce qui indique que ces fleurs sont autocompatibles, capables de s’autoféconder en absence de visites.
Cependant, les fruits issus de fleurs pollinisées par des insectes renferment en moyenne 18 graines/fruit contre seulement 7 sur les fleurs isolées des insectes. L’érythrone a donc fortement besoin des insectes pollinisateurs pour assurer une descendance abondante. On a observé que les fleurs plus petites produisaient moins de graines par fruit que les plus grandes justement à cause du moindre nombre de visiteurs sur les premières, moins attractives.
Fourmis
Les fleurs passent très vite et les tépales sèchent et tombent, exposant l’ovaire et ses ovules fécondés. Celui-ci gonfle et se transforme en une capsule globuleuse à trois angles peu marqués ; elle est penchée vers le bas comme la fleur initiale. Ainsi, arrivée à maturité, quand elle s’ouvre (fruit sec déhiscent), les graines tombent au sol. Dans une étude menée en Espagne, 99% des graines tombent dans un rayon de 20cm en moyenne autour du pied mère mais avec de fortes variations d’un pied à l’autre. C’est la dispersion primaire peu efficace donc en termes de déplacements.

Fleur fanée 
Jeune fruit vert 
Le style reste sur l’ovaire gonflé 
Fruit sec trigone (Cliché C. Berg ; C.C. 4.0)
Mais ces graines portent un appendice huileux, un élaïosome assez gros, très convoité par certaines espèces de fourmis (voir l’exemple de l’euphorbe épurge) ; elles s’emparent de la graine en la saisissant par cet appendice et la traînent vers leur fourmilière ; avant d’entrer, elles détachent la graine rejetée à l’extérieur et emportent l’appendice dans la colonie comme nourriture pour le couvain. Ainsi, les graines peuvent parcourir une certaine distance (de l’ordre de quelques mètres en général) et se retrouvent souvent dans des sites assez favorables ensuite pour leur germination. On parle de dispersion secondaire et de myrmécochorie (myrmeco, fourmi ; chori : transport).
Cette dispersion secondaire par les fourmis varie beaucoup selon les habitats ; sur certains sites, elle est nulle et sur d’autres elle concerne de 12 à 23% des graines au sol. En forêt, c’est souvent l’espèce Formica lugubris qui intervient. Les graines fraîchement tombées ont beaucoup plus de chances d’être emportées car ensuite l’élaïosome se dessèche et l’attractivité s’amoindrit. On a aussi observé que les graines n’étaient pas consommées par les rongeurs (mulots) qui sont souvent des prédateurs redoutables des graines.
Au moment de la dispersion, les embryons dans les graines sont encore indifférenciés. La germination commence en hiver avec l’émergence de la radicule ; les cotylédons et premières feuilles sortiront en fin d’hiver.
Dent-de-chien

La hampe florale haute de 10 à 20cm émerge entre deux feuilles opposées au ras du sol ; elles sont produites depuis un bulbe blanc, mince et allongé, habillé d’une tunique membraneuse. C’est lui qui a donné ce qualificatif de dent-de-chien (dens-canis) à cause de la ressemblance avec un croc de chien. Ce nom est repris dans plusieurs langues : dog’s tooth en anglais, dente di cane en italien
Grâce à ce bulbe chargé de réserves nutritives, la plante va pouvoir émerger tôt, produire feuilles et fleur très vite ; elle profite de l’absence de feuillage dans les canopées : avec un plein éclairage, elle effectue une photosynthèse très active qui permet de recharger les réserves du bulbe pour l’année suivante. En moins de deux mois, le cycle est bouclé et la plante disparaît, au moins pour ses parties aériennes : on parle d’éphémère vernale pour un tel cycle si rapide.
Le bulbe mature élabore de jeunes bulbes secondaires ou caïeux (comme les « gousses » d’ail) qui assurent une multiplication asexuée. Ceci explique pourquoi souvent un pied donné est accompagné d’autres feuilles visiblement plus jeunes émises par ces caïeux.

On a étudié individu par individu une population dans le nord des Apennins italiens. Sur 590 érythrones suivies, 19 seulement ont fleuri, 442 étaient matures mais n’ont pas fleuri et 130 étaient à un stade juvénile avec une seule feuille. Les pieds fleuris émergent dès fin février alors que les seconds le font à mi-mars et les derniers fin mars. Selon leur âge, les feuilles produites sont différentes (voir ci-dessous). La phase de maturation du bulbe depuis le stade plantule semble être très logue et nécessaire pour produire un bulbe assez grand pour permettre de réaliser le cycle complet en mode éclair. Pendant ce temps, progressivement, le bulbe en formation s’enfonce chaque année un peu plus.
Deux feuilles
Longuement pétiolées, de forme elliptique à lancéolée, les deux feuilles s’étalent de chaque côté et attirent l’œil par leur coloration originale : sur un fond vert bleuté (glauque), des taches arrondies brun rougeâtre ou parfois presque blanches et, plus discrètes, de taches diffuses argentées. L’ensemble forme un motif bariolé très élégant.


Dans la population étudiée dans les Apennins (voir ci-dessus), on a observé trois types majeurs de feuilles selon les individus : des feuilles fortement argentée (62 à 74%), des argentées et vertes (23 à 32%) et des entièrement vertes (3-5%). Une fois le stade juvénile passé, un même individu produit toujours le même type de feuille ce qui suggère un déterminisme génétique.

Certains pieds n’ont qu’une feuille mais alors très grande en forme de bouclier arrondi : ce sont des plantes n’ayant pas atteint le stade de la maturité florale qui ne fleurissent pas encore. De même, les individus juvéniles n’ont qu’une petite feuille oblongue à lancéolée, avec un motif de taches brun clair peu marqué ou bien entièrement vertes sans motif.
On retrouve de telles feuilles tachetées chez diverses espèces d’érythrones nord-américaines connues là-bas sous le surnom de trout-lily, soit lis-truite en référence à la coloration des truites de rivière : une comparaison qui pourrait presque convenir aussi à notre érythrone.
Les taches brunes s’effacent progressivement à partir du moment où la fleur passe et virent graduellement au vert luisant. Autrement dit, l’aspect tacheté n’est que temporaire.
Taches

Dessous : les taches sont à peine visibles car localisées sous l’épiderme supérieur
Les taches brunes se forment sous l’épiderme dans la couche supérieure du tissu chlorophyllien interne (mésophylle) : des pigments solubles ou anthocyanes s’accumulent dans les vacuoles des cellules. Leur position explique qu’elles paraissent un peu estompées vues de l’extérieur compte tenu de l’épiderme qui s’interpose.
Les taches argentées, quant à elles, proviennent d’un décollement de l’épiderme par rapport au tissu sous-jacent ce qui induit une réflexion de la lumière au niveau des minces espaces ainsi libérés et remplis d’air ; il s’agit donc d’une couleur physique.
Une analyse précise des différents tissus tant au niveau des taches qu’en dehors montre qu’elles ne diffèrent pas du tout en termes de pigments photosynthétiques ; donc, la présence de ces taches ne diminue pas le potentiel photosynthétique des feuilles, contrairement à ce qu’on pourrait penser. Or leur efficacité est cruciale pour que la plante puisse reconstituer ses réserves souterraines pendant la courte période de végétation dont elle dispose.
Le verdissement des taches brunes (voir ci-dessus) résulte de la résorption des anthocyanes dans les vacuoles ; il est d’ailleurs plus rapide chez les individus non fleuris par rapport aux fleuris.
Tenue léopard !
Trois hypothèses ont été avancées quant à la fonction de ces feuilles tachées : attirer les pollinisateurs ; protéger les tissus foliaires d’une trop forte intensité de lumière ; procurer un camouflage vis-à-vis des vertébrés herbivores. L’effacement progressif des taches brunes invalide les deux premières hypothèses ; par contre, il se pourrait que les taches argentées jouent un rôle attractif envers les pollinisateurs au moment de la floraison permettant un repérage des plantes de loin.
L’hypothèse du camouflage semble bien la plus plausible, mais non validée expérimentalement. On l’a avancé à partir d’un constat : diverses plantes forestières herbacées basses, dont le feuillage passe l’hiver ou émerge tôt au printemps comme l’érythrone, présentent de tels motifs tachetés ou marbrés sur leurs feuilles : cirons l’arum d’Italie (feuilles marbrées de blanc ivoire), le cyclamen à feuilles de lierre (marbrures claires), les pulmonaires ou le lamier jaune aux taches argentées.

Arum d’Italie 
Lamier jaune 
Cyclamen à feuilles de lierre 
Pulmonaire
Par ailleurs, en Amérique du Nord, le genre Erythronium est très diversifié et toutes les espèces partagent un cycle court au printemps. Sur seize espèces étudiées, les trois du type éphémère vernale (voir ci-dessus) ont des feuilles tachées ; les treize autres ont soit des feuilles tachées ou pas : les espèces non tachées vivent à haute altitude dans des prairies alpines très ouvertes ; les tachées poussent au contraire dans des sites plus fermés du type maquis ou bois clairs.
Ces plantes sont effectivement très exposées à des risques de prédation de leur feuillage car leurs feuilles sont très riches en azote, ayant été produites à partir des réserves des bulbes. On pense que ces taches cassent la silhouette des feuilles telles qu’elles peuvent être perçues par des vertébrés dépourvus de la vision des couleurs (cas des mammifères herbivores) dans les sous-bois éclairés au printemps par des taches de lumière. En plus, les grands herbivores, à cette période de l’année, se concentrent sur les jeunes feuillages frais émergents plus appétents.
Pour la survie de l’érythrone, conserver ses feuilles intactes le temps de son cycle est crucial ; sans elles, ou si elles sont trop réduites, elle ne peut reconstituer les réserves du bulbe pour la saison suivante ; or, nous avons vu que l’élaboration de ce bulbe avait demandé une longue période de plusieurs années, soit un investissement considérable en termes énergétiques.
Néanmoins, on observe une assez forte prédation de la part des limaces mais avec un impact plus limité (feuilles trouées).
Localisée
En France, si l’érythrone dent-de-chien peut être très abondante dans ses stations, elle reste globalement assez rare du fait notamment de son aire de répartition assez limitée et de ses exigences écologiques assez étroites.
On la trouve depuis 200m (comme dans le Sud-Ouest) jusqu’à 2300m dans l’étage alpin. Son aire disjointe couvre le sud du Jura, les Préalpes, les Alpes-Maritimes, le quart sud-ouest du Massif Central d’où elle rejoint la chaîne pyrénéenne jusque dans les plaines du pays Basque.
En Europe, son aire se limite à une grande moitié sud : Espagne (dont la Chaîne Cantabrique), France donc, Nord Italie, péninsule des Balkans en descendant vers la Grèce ; quelques populations à l’Est des Balkans.

Prairie en bordure d’une rivière (Auvergne) 
En sous-bois (cliché Huhulenik ; C.C. 3.0.)
En région montagneuse, l’érythrone recherche des sites assez abrités comme des vallons ou des bas de pentes. Elle habite des milieux boisés clairs (chênaies, charmaies, hêtraies, châtaigneraies) ou dans des milieux plus ouverts comme des prairies en bord de rivière, des landes herbeuses ou des pelouses alpines. Elle se comporte en espèce héliophile (lumière) tolérant la demi-ombre. Elle est sensible aux gelées tardives du fait de son cycle printanier. Son aire globale apparaît effectivement limitée d’une part par des hivers trop froids (absente d’une bonne partie des Alpes) et des saisons estivales sèches prolongées (milieu méditerranéen).
En Europe centrale (où elle est très rare), on a noté sa plus grande abondance dans des peuplements forestiers plus anciens ; une densité élevée de jeunes arbres lui est défavorable. Les forêts de résineux sont pratiquement évitées. Évidemment, compte tenu de son caractère vivace, l’exploitation forestière intensive le défavorise nettement.
On a observé sur d’autres espèces (Amérique du nord) que la hausse moyenne des températures des sols tendait à faire diminuer la taille des bulbes ce qui laisse planer une ombre sur l’avenir de cette espèce : des bulbes plus petits signifient un moindre succès reproductif. On a aussi observé des décalages de calendrier d’émergence entre ces plantes et leurs pollinisateurs pouvant là aussi induire une baisse de la reproduction (voir la pollinisation).
Côté sols, elle a besoin d’une certaine humidité (espèce mésophile), d’une richesse en nutriments favorable à son développement rapide et court, sur des sols argileux souvent riches en calcaire (carbonatés). Cependant, dans le sud-ouest, elle se comporte en espèce nettement acidiphile dans les landes atlantiques et les boisements de châtaigniers.
Parentés
Le genre Erythronium renferme entre 25 et 27 espèces qui partagent plusieurs caractères : un bulbe allongé et ovale, une paire de feuilles basales, une ou plusieurs fleurs au sommet d’une hampe florale nue, un cycle court centré sur le printemps. Les variations entre espèces portent sur la couleur des fleurs (avec par exemple des fleurs jaunes), le nombre de fleurs par hampe, la présence ou pas de taches sur les feuilles et la capacité de multiplication végétative.

E. oregonum (cliché C.C. 2.5) ; Ouest des USA 
E. pusaterii (cliché J. Game ; C.C. 2.0) ; hampe à plusieurs fleurs ; endémique d’un comté californien 
E. umbilicatum (cliché D. Goldman ; C.C. 4.0) ; Sud-Est des USA 
E. helenae (cliché Dinkum ; C.C. 3.0) ; feuilles non tachées ; endémique Californie
Une analyse génétique a permis d’identifier trois grands groupes géographiques ayant évolué indépendamment :
- eurasiatique de 4 espèces : E. dens-canis, E. caucasicum, E. sibiricum et E. japonicum
- dans l’Est de l’Amérique du nord
- dans l’Ouest notamment dans les montagnes de Californie.

L’Amérique du nord héberge la majorité des espèces et elles s’hybrident facilement entre elles ce qui confirme leur évolution récente (Pléistocène).
Au sein de la famille des Liliacées, dans la grande sous-famille des Lilioidées (Fritillaires, Lis, …), les érythrones sont étroitement apparentées aux Tulipes (Tulipa) et à un autre genre proche de 5 espèces asiatiques, Amana. On les réunit avec les Gagées (Gagea) dans une tribu à part (les Tulipées) qui se détache à la base de la sous-famille : autrement dit, érythrones, tulipes et gagées sont d’origine plus ancienne que lis et fritillaires.
Pour autant, à cause de la structure « renversée » de la fleur, on confond souvent l’érythrone avec d’autres fleurs non apparentées : ainsi, la ressemblance est frappante avec les fleurs des cyclamens et surtout celles d’un genre nord-américain, cultivé comme ornementale, les gyroselles (Dodecatheon). Ces deux genres de plantes se classent dans une tout autre famille très éloignée dans l’arbre des parentés : les primulacées. Il s’agit d’une simple convergence quant à la forme générale des fleurs ; ainsi par exemple, cyclamens et gyroselles ont des fleurs à 5 pétales et non pas à 6 tépales ! Les feuilles sont aussi très différentes car les érythrones sont des Monocotylédones (feuilles à nervures parallèles) alors que les autres sont des Dicotylédones (nervures ramifiées).
Interactions
En Sibérie, l’espèce proche E. sibiricum faisait l’objet d’une exploitation humaine par les ethnies locales au moins jusqu’au 18ème siècle : on récoltait les bulbes, mangés avec du renne ou lait de vache. Leur richesse en amidon et leur non toxicité en faisaient une ressource complémentaire non importante. La récolte avait lieu au printemps juste à l’émergence et on savait aussi les chercher dans les réserves des terriers de petits rongeurs lors d’épisodes froids. Un commerce local important existait autour de cette ressource ! Les feuilles, comestibles, étaient aussi consommées cuites à l’eau. De même au Canada, d’autres espèces étaient récoltées par des ethnies amérindiennes : ils avaient sans doute observé que les ours déterraient les bulbes avec avidité pour les consommer.
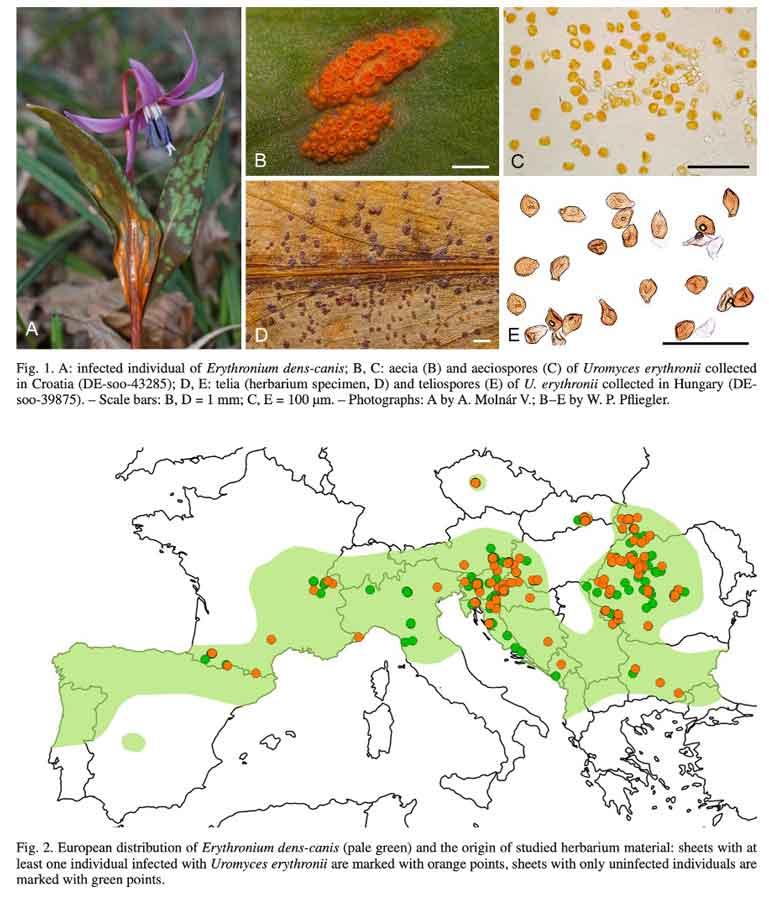
A part les herbivores (voir les feuilles tachées), on ne connaît que très peu d’espèces interagissant avec l’érythrone. Parmi les parasites, il y a une rouille spécifique Uromyces erythronii qui effectue tout son cycle sur cette plante. Elle se manifeste entre autres par de larges plaques orangées pustuleuses sous les feuilles. Une étude sur des échantillons d’herbier a montré que le parasite se rencontrait sur toute l’aire de répartition européenne. Le taux d’infection, faible, semble être resté inchangé depuis 200 ans.
Transgénique naturelle !
Nous allons terminer ce long tour d’horizon sur cette plante décidément étonnante par un « scoop » inattendu révélé en 2022. En analysant un petit secteur de l’ADN (porté par les chromosomes) des érythrones, les chercheurs ont eu la surprise d’y découvrir une portion assez importante qu’ils ont identifié comme provenant de l’ADN d’une autre plante, une potentille (Potentilla). Or, les potentilles sont très éloignées en termes de parenté des érythrones : ce sont des Dicotylédones Rosacées versus Monocotylédones Liliacées ; pour bien comprendre, c’est comme si dans notre ADN d’Humain on trouvait une séquence sur un de nos chromosomes provenant d’un mammifère très éloigné de nous en parentés : disons par exemple un tatou ! Autrement dit, les érythrones sont quelque part des organismes transgéniques naturels !
La présence de cette séquence « étrangère » sur un des chromosomes des érythrones signifie qu’il y a eu à un moment de l’histoire évolutive de ces plantes un Transfert Horizontal de Gènes : un échange de gènes qui franchit la barrière des espèces (d’où le qualificatif d’horizontal) et non pas comme classiquement d’une génération à l’autre entre individus de la même espèce (vertical). L’ADN ainsi transféré a conservé sa fonctionnalité : il code bien des protéines mais moins efficacement semble-t-il que dans son hôte originel (potentille).
Un tel transfert est répandu chez les organismes unicellulaires comme des bactéries mais rarissime chez des organismes pluricellulaires même si on commence à en découvrir de plus en plus : pas facile de les détecter ! Ainsi, on a démontré leur fréquence entre espèces de graminées : mais là on reste entre espèces de la même famille. Ici, la barrière franchie est considérable.
On a pu établir que ce transfert avait eu lieu au niveau de l’ancêtre commun entre l’E. dent-de-chien et l’E. du Caucase, assez proche (voir ci-dessus), soit il y a plusieurs millions d’années.
Quelles conséquences pour les érythrones ? Difficile à savoir : cela a pu aboutir à des innovations comme l’adaptation à un nouvel environnement ou une plus grande plasticité écologique, ou un changement dans le métabolisme, ou ….
Reste une question qui brûle les lèvres : comment cela s’est-il passé ? L’hypothèse la plus plausible (mais il y en a d’autres) serait par le truchement de piqûres de puceron ayant absorbé de la sève sur des potentilles avant d’aller sur des éryhtrones. Dans cette sève, il y avait un virus végétal (très courants) qui avait incorporé un brin d’ADN des potentilles (ils savent très bien faire). Inoculé ensuite dans l’érythrone, il aurait transféré ce brin sur l’ADN de l’érythrone. C’est d’ailleurs une des techniques utilisées pour fabriquer des OGM.
Bien, ce sera tout pour l’instant : sacrée plante quand même ! Et il y a sans doute encore bien d’autres choses à découvrir … comme pour de nombreuses autres plantes. Le vertige de la découverte infinie du Vivant : largement de quoi occuper plusieurs vies !
Bibliographie
Guide expert des Plantes de montagne. F. Le Driant et al. Ed. Biotope 2022
Population structure of Erythronium dens-canis L. (Liliaceae) in the northern Apennines (Italy). Italian Botanist 4: 1–14. Pupillo P, Astuti G (2017)
Erythronium dens-canis L. (Liliaceae): an unusual case of change of leaf mottling Nicoletta La Rocca et al. Plant Physiol Biochem. 2014 :74:108-17
Leaf Mottling: Relation to Growth Form and Leaf Phenology and Possible Role as Camouflage T. J. Givnish Functional Ecology, Vol. 4, No. 4 (1990), pp. 463-474
Seed dispersal in Erythronium dens-canis L. (Liliaceae): variation among habitats in a myrmecochorous plant Pablo Guitián Plant Ecology 169: 171–177, 2003.
Effect of structure and dynamics of forests on the occurrence of Erythronium dens-canis Stanislav Vacek Journal of Forest Science, 66, 2020 (9): 349–360
Gathering dog’s tooth violet (Erythronium sibiricum) in Siberia Sabira STÅHLBERG & Ingvar SVANBERG
Distribution, infection rates and DNA barcoding of Uromyces erythronii (Pucciniaceae), a parasite of Erythronium (Liliaceae) in Europe TIMEA NAGY Willdenowia Annals of the Botanic Garden and Botanical Museum Berlin 2018
Phylogeny, Age, and Evolution of Tribe Lilieae (Liliaceae) Based on Whole Plastid Genomes. Li J et al; (2022) Front. Plant Sci. 12:699226.
Intact ribosomal DNA arrays of Potentilla origin detected in Erythronium nucleus suggest recent eudicot-to-monocot horizontal transfer Laszlo Bartha et al. New Phytologist (2022) 235: 1246–1259
