Les antennes des insectes captent la 5S
10/02/2022 Que les geeks se calment, il ne s’agit pas d’une nouvelle génération de téléphonie mobile révolutionnaire : 5S n’est qu’un jeu de mots raccourci pour signifier 5 Sens ! En effet, les antennes des insectes sont des organes sensoriels (entre autres) capables de percevoir au moins cinq sens dont plusieurs nous sont complètement étrangers. Nous allons donc d’abord découvrir comment sont faites les antennes, puis leurs différentes fonctions sensorielles et nous nous interrogerons sur l’origine évolutive de ces organes et quels autres animaux en dehors insectes en sont dotés.
Articulées

Où chercher les antennes et comment ne pas les confondre avec d’autres organes proches d’aspect ? Appendices formés d’articles, elles sont toujours disposées en une paire insérée sur la capsule rigide de la tête, sur le « front », donc au-dessus des pièces buccales. Ces dernières, toutes proches, présentent souvent elles aussi des pièces allongées articulées : des palpes qui accompagnent les mandibules et les maxilles ; mais il se trouvent autour de la bouche et donc « sous » la tête.

De même, il peut y avoir au bout de l’abdomen des appendices articulés, des cerques : ce ne sont pas non plus des « antennes postérieures » !

Cerques en filets au bout de l’abdomen : éphémère 
Cerques courts et pointus d’une blatte 
Deux cerques très courts de chaque côté de l’énorme ovipositeur d’une éphippigère
Chez certains insectes, la tête peut aussi porter des ornementations allongées, parfois ramifiées : des cornes comme chez de nombreux scarabées. Il s’agit soit de pièces buccales transformées (comme les mandibules des lucanes : voir la chronique) et dans ce cas sont donc par paire, soit d’expansions de cuticule (la peau durcie des insectes) souvent impaires.

Dans tous les cas, elles ne sont jamais composées de segments articulées et donc d’une seule pièce. Enfin, chez les larves de la majorité des espèces à métamorphoses complètes, les antennes sont souvent bien plus réduites (voire quasi absentes) que chez les adultes.

Larves de doryphores 
Larve : antenne réduite à un moignon 
Tête de doryphore adulte : antenne à articles nombreux
Le mot antenne est passé dans le langage usuel pour désigner les tiges émettrices des appareils radio mais ce même mot appliqué aux insectes remonte lui bien avant l’apparition des premières « antennes radio ». On a commencé à l’utiliser en entomologie (la science qui étudie les insectes) à partir du début du 18èmesiècle par analogie avec une pièce de bateau : une longue vergue soutenant une voile et connue depuis au moins le 13ème siècle sous l’appellation de antaine.
Tripartites

En dépit des infinies variations de taille et de forme, les antennes des insectes conservent une remarquable unité de structure avec trois segments principaux. La pièce basale de l’antenne ou scape (de scapus, tige) s’articule sur une membrane dans un creux entouré d’une structure durcie en forme d’anneau (torulus) sur le dessus de la tête ; mais l’antenne n’est pas libre sur cette membrane : elle pivote sur une protubérance rigide émanant du bord du torulus (antennifère). Grâce à cette articulation, l’insecte peut bouger son antenne en bloc, dans un plan vertical, via des muscles internes rattachés au scape.

Sur le scape, qui est en général le segment le plus robuste de l’antenne, se raccorde un second segment, le pédicelle. Il porte à son extrémité distale un organe sensoriel très important, sensible aux mouvements du reste de l’antenne, l’organe de Johnston (voir ci-dessous : audition). Le pédicelle est articulé avec l’extrémité supérieure du scape et là aussi ses mouvements sont contrôlés par des muscles internes entre le scape et lui.

Base des antennes d’un « capricorne » : Morime rugueux ; noter le torulus basal en forme d’anneau 
Antennes et palpes buccaux d’un perce-oreille (Dermaptère)
Enfin, le pédicelle porte le flagelle souvent en forme de filament et composé d’articles, les flagellomères. Le flagelle commence donc à partir du troisième article en partant de la tête, après le scape et le pédicelle. Le nombre de ces articles varie considérablement selon les espèces et sert souvent de critère d’indentification : primitivement, il était de huit ; il peut être très réduit chez certains groupes comme les libellules (Odonates) où il se compose d’un article.

Antennes très simplifiées d’une libellule (Odonate) 
Demoiselle (Odonate)
Ces articles sont articulés entre eux mais ils ne renferment pas de muscles internes si bien que le flagelle ne bouge que d’un bloc au niveau de l’articulation avec le pédicelle. Cependant, chez les Scarabéidés (Coléoptères : hannetons, cétoines, …), le flagelle se compose de lamelles repliées au repos mais que l’insecte peut déployer pour capter des odeurs via des changements de pression du liquide interne et l’élasticité des parois et des membranes articulaires.

Antenne lamellée d’un hanneton foulon (Coléoptère Scarabéidé) 
Collembole (Hexapode non-insecte)
Chez les Hexapodes non-insectes (voir le paragraphe final ) tels que les collemboles ou les diploures, les flagelles des antennes possèdent encore des muscles internes. Chez divers groupes d’insectes des lignées basales (les plus anciennes), on observe souvent une fragmentation des articles du flagelle en sous-unités ou anneaux très nombreux comme chez les lépismes (Thysanoures), les archéognathes ou les blattes et mantes.

Archéognathe 
Blatte 
Mante
Chez de nombreux insectes dont les Coléoptères (voir la chronique) et les Hyménoptères (voir la chronique), l’extrémité supérieure du pédicelle s’articule avec le flagelle de sorte qu’il peut se replier à angle droit : on parle d’antennes géniculées (de genou).


Fourmis en train de palper des pucerons 
Poliste butinant des fleurs de fenouil
Ce coude facile à observer permet par exemple aux fourmis (Hyménoptères) de suivre les pistes odorantes laissées par les congénères ou de tapoter les pucerons avant la traite du miellat ou aux abeilles et guêpes de « renifler » les fleurs visitées ; chez les scarabées et les charançons (Coléoptères), le coude permet de plier l’antenne notamment quand ils adoptent une « position du mort » en repliant leurs pattes comme moyen d’échapper à un prédateur.

Antennes géniculées d’un charançon (Liparus) 
Charançon sp : les antennes peuvent se replier sur les côtés dans une gouttière 
Lixus (charançon) et ses antennes géniculées
On observe aussi souvent une dilatation en massue de la partie terminale comme chez les mini-guêpes Chalcidiens (responsables de galles : voir la chronique) ou certains coléoptères : on appelle alors funicule la partie entre la massue et le pédicelle porteur.

Variées

Compte tenu de l’importance cruciale des antennes comme organes multi sensoriels (voir ci-dessous) et notamment comme outils de communication sexuelle (via la chémoréception des phéromones), elles ont subi au cours de l’évolution de très fortes pressions de sélection naturelle (dont la sélection sexuelle) qui ont conduit à une extrême diversification au niveau des formes et des tailles. Nous allons en présenter quelques types répandus mais sans être exhaustif vu l’étendue de cette diversité morphologique.

Antennes sétacées des phryganes 
Antennes de longicorne : le grand capricorne 
Chez les punaises, le flagelle des antennes a peu d’articles : Corée marginée 
Punaise américaine des pins
Chez un grand nombre d’espèces, les antennes sont classiquement filiformes : simples et droites avec de multiples articles. Une variante dite sétacée (dérivé de seta pour soie) se rétrécit de la base vers l’extrémité comme chez les blattes, les éphémères, les perles ou les phryganes. Chez divers coléoptères ou chez les termites les articles arrondis donnent à l’antenne l’apparence d’un collier de perles : on parle d’antenne moniliforme (de monile, collier) ; cet adjectif comme d’autres qui suivent se retrouve en botanique pour décrire par exemple les filets des étamines : on ne va pas parler de convergence pour autant ! Dans la chronique sur les cardinaux, ces coléoptères rouges, nous avons présenté les deux types d’antennes dentelés selon les sexes : serriforme avec un bord en dents de scie ou pectiniforme (pecten = peigne) avec un bord en peigne ; ce dernier type se retrouve chez des taupins (Élatéridés : voir la chronique) ou des tenthrèdes (Hyménoptères).

Taupin pourpre aux antennes fortement pectinées 
Détail d’une antenne 
Antenne pectinée de cardinal adulte 
La larve a des antennes « normales »
Chez de nombreux papillons de jour et des coléoptères, les derniers articles du flagelle s’élargissent et forment ensemble une massue soit de manière brutale soit de manière progressive (antenne clavée ou claviforme ; de clava, massue ; voir les champignons nommés clavaires en forme de massue). Nous avons ci-dessus évoqué les étranges antennes lamelliformes des scarabéidés à articles en lamelles ; quand elles sont au repos, on a l’impression d’une grosse masse aplatie un peu courbée. D’ailleurs autrefois on nommait ces coléoptères des Lamellicornes. Dans le même style, on nommait longicornes tous les coléoptères aux longues antennes souvent tenues rabattues vers l’arrière et typiques de la famille des Cérambycidés (voir la chronique sur le morime par exemple).

Antennes courtes des criquets par rapport aux longues antennes des sauterelles 
Antennes filiformes chez de nombreux microlépidoptères nocturnes comme ce Ptérophore. 
Antennes en massue progressive chez ce macroglosse (Sphingidé) 
Antenne en club de golf du machaon (Papillon de jour)
Chez les diptères, on oppose deux grands groupes sur la base d’antennes radicalement différentes. Les Nématocères (moustiques, tipules et « moucherons ») ont des antennes filiformes (de nematos filament et keros, corne) « classiques » avec de nombreux articles de même taille. Par contre, les Brachycères qui regroupent les mouches dont les syrphes, les taons, les bombyles (voir la chronique), possèdent des antennes très réduites (brachy = court) à trois articles : le scape, le pédicelle et un flagelle en forme de massue arrondie portant un long poil ramifié ou arista ; on parle d’antenne aristée.

Antenne plumeuse de chironome (Diptère Nématocère) 
Antennes filiformes simples de tipule (Nématocère) 
Antenne de syrphe : 3 articles et une soie sur le flagelle en massue (Brachycère) 
Antennes de syrphe vues par dessous 
Antennes à soie plumeuse d’une volucelle (syrphe) 
Arista plumeuse d’une antenne de mouche
Enfin, on ne peut manquer les belles antennes plumeuses ramifiées typiques des mâles de certains papillons de nuit (dont les saturnidés ou les lasiocampidés) et des diptères nématocères comme les antennes des moustiques ou des chironomes. Nous reparlerons beaucoup de ce type d’antenne spécialisé dans la captation des molécules odorantes.


Mâle de bombyx disparate 
Gros plan sur les antennes plumeuses
Sensorielles
Le succès évolutif considérable des insectes s’appuie en partie sur la capacité de leurs sens à interpréter leur environnement et à en discriminer des éléments à une échelle très fine ; cette capacité sensorielle globale a notamment permis le développement du vol (vision en vol, perception de l’orientation, …). La gamme des sens dont disposent les insectes est bien plus étendue que la nôtre : si la vision est bien développée, l’ouïe et l’olfaction dominent dans leur perception de l’environnement et surtout dans la communication entre individus et dans les interactions. Contrairement à nos organes sensoriels pour la plupart centralisés et plus ou moins internes, les insectes déploient à la surface de leur cuticule (la peau durcie) une multitude de micro-récepteurs sensoriels, des sensilles, chargées de détecter les stimuli externes, de les transcoder et les transmettre au système nerveux central via des fibres nerveuses. Ces sensilles peuvent prendre différentes formes selon les fonctions et les emplacements : des poils, des creux, des protubérances, … Dans ce contexte, les antennes concentrent à leur surface un grand nombre de sensilles de différentes catégories capables de percevoir a minima cinq grands types de stimuli externes : les déformations et mouvements, les sons ou vibrations, la chaleur, l’humidité et les odeurs !

Poils sensoriels sur des antennes d’un carabidé 
Poils sur les articles du flagelle 
Lamelles d’antenne de hanneton et leurs sensilles
Chacune de ces sensilles n’est formée que de quelques cellules associées dont une cellule nerveuse. Quand par exemple, une sensille olfactive d’une antenne est touchée par des molécules odorantes dans le flux d’air qui circule autour de l’insecte, la cellule nerveuse, en lien avec la partie émergée qui capte les odeurs, s’active et envoie un train de potentiels d’action (signaux électriques) vers un lobe du système nerveux central qui renvoie ensuite l’information vers deux corps pédonculés associés qui l’analysent et interprètent l’odeur. En laboratoire, on est capables d’enregistrer et de suivre la somme de tous ces potentiels d’action émis par les différentes sensilles suite à une stimulation expérimentale (par exemple, envoyer sur l’antenne un flux odorant) à l’échelle de l’antenne ; on obtient ainsi un enregistrement appelé électro-antennogramme. Grâce à cette technique de micromanipulation on a beaucoup appris sur le fonctionnement sensoriel des antennes.
Parcourons donc les différentes capacités sensorielles assurées par les antennes
Mouvements et position
Le corps des insectes est entièrement enveloppé d’une peau plus ou moins rigide (exosquelette), parfois très durcie en carapace comme chez les Coléoptères (voir la chronique) ; pour autant, elle reste soumise à des distorsions imposées par les mouvements de l’environnement ambiant ainsi que ceux résultant des contractions des muscles internes. L’insecte se trouve ainsi en permanence soumis à toutes sortes de stimuli mécaniques : toucher, gravité, étirement du corps, position, pressions et vibrations liées aux mouvements de l’air et/ou du substrat de déplacement. Pour capter ces informations cruciales, les insectes disposent de sensilles dites trichoïdes (i.e. en forme de poils) réparties à la surface du corps et qui émergent depuis de légers creux dans la cuticule. Elles assurent donc la fonction tactile mais bien plus : ainsi, certaines de ces sensilles réagissent aux vibrations de l’air engendrées par des bruits ! Par ailleurs, il existe d’autres sensilles (d’au moins trois types différents) spécialisées dans la perception de la position relative du corps et son orientation par rapport à la gravité : on parle de propriocepteurs. Elles sont concentrées autour du « cou » (articulation entre la tête et le thorax) et aux articulations des pattes et des ailes avec le corps.


Haltères jaunes chez ces Tipulidés 
Haltères brunes d’une syrphe
On se doute que ces perceptions prennent toute leur importance en vol, souvent très rapide et avec une grande manœuvrabilité. Chez les Diptères tels que les mouches ou les moustiques et tipules, les ailes postérieures se sont transformées en organes en forme d’antenne avec une massue terminale : les haltères, placées sur les côtés du thorax, juste en arrière et en contrebas des ailes antérieures. Ces haltères, par leurs mouvements giratoires, détectent les « forces » de Coriolis liées au déplacement dans l’air et assurent ainsi la stabilité en vol en permettant d’ajuster la position du corps. Les papillons de nuit par contre ont, comme la majorité des insectes, deux paires d’ailes et n’ont pas de telles haltères. On a montré que leurs antennes vibrent en vol ce qui les rend capables de percevoir les forces de Coriolis pendant leurs déplacements : les vibrations des antennes sont transcodées par des organes situés sur le pédicelle des antennes (organe de Johnston : voir ci-dessous) en messages nerveux dont la fréquence correspond à la position relative dans l’air ; expérimentalement, si on enlève le flagelle des antennes, le papillon perd sa stabilité en vol.
Audition

Ce terme hérité de notre perception sensorielle traduit en fait bien mal celle des insectes capables de percevoir des « sons » ou vibrations dans des gammes de fréquences bien au-delà de notre bande passante. Il vaudrait mieux parler de perception de vibrations sonores qui intervient dans la communication entre individus au sein d’une espèce (communication acoustique) avec notamment la production de « chants » comme chez les Orthoptères (voir la chronique sur la sauterelle ponctuée) ou dans la détection des prédateurs comme la capacité des papillons de nuit à percevoir les ultra-sons émis par les chauves-souris en vol. Cette perception sonore va de la réception des vibrations du sol ou du substrat sur lequel vivent les insectes jusqu’à capter des sons lointains. Divers organes sensoriels sont impliqués dans ce registre : les organes spécialisés comme des membranes cuticulaires ou tympans bien connus chez les Orthoptères (voir la chronique), des sensilles trichoïdes réparties sur le corps ou les cerques abdominaux des grillons ou des blattes sensibles au moindre courant d’air.
Les antennes portent quant à elles des structures spécialisées dans la réception des vibrations : les organes chordotonaux. Chacun d’eux se compose d’une à plusieurs unités, des scolopidies complexes, formées de trois cellules alignées et implantées sur la cuticule. Tous les insectes adultes et de nombreuses larves possèdent au niveau du pédicelle un organe chordotonal particulier, l’organe de Johnston dont la fonction première est de détecter les mouvements du flagelle de l’antenne par apport au pédicelle (voir ci-dessus) comme pour apprécier la vitesse de l’air en vol. Mais secondairement, cet organe sert aussi à « entendre ». Chez les mâles de moustiques et de chironomes, le pédicelle renflé des antennes renferme de nombreuses scolopidies fixées sur sa paroi et sur celle du premier article du flagelle au-dessus. Cet organe de Johnston modifié permet aux mâles de repérer la tonalité du son émis par les vibrations des ailes des femelles en vol. On a souvent surnommé cet organe « oreille de l’abeille » car on l’a découvert chez ces dernières en lien notamment avec la danse frétillante qu’effectuent les ouvrières de retour à la ruche.
Autres sens
Chez une fourmi tropicale migratrice, on a mis en évidence la présence de composés ferreux et de silicates à l’intérieur des tissus de l’organe de Johnston ; ils seraient extraits du sol et incorporés. Les propriétés magnétiques de ces microparticules fourniraient à ces fourmis un sens magnétique permettant l’orientation. Il existe aussi au niveau des antennes des récepteurs à la chaleur mais peu nombreux soit exposés soit dans des creux et associés dans la même sensille à des récepteurs à l’humidité.
Mais le sens le plus extraordinairement développé et localisé dans les antennes des insectes reste l’olfaction ou la perception de molécules chimiques odorantes (chémoréception). Ce sens est bien plus développé que chez les vertébrés. La position en avant des antennes et leur déploiement les met le plus souvent en première position pour entrer en contact avec des molécules chimiques transportées par l’air. Devant l’étendue et la complexité du rôle des antennes dans ce sens chez les insectes, nous lui consacrerons une chronique à part entière car il a des implications multiples dans la vie des insectes notamment au niveau de la reproduction. Ici, nous ne ferons que décrire brièvement les sensilles olfactives des antennes.
Les sensilles olfactives des antennes sont de deux grands types. Certaines ne possèdent qu’un seul pore terminal ou central (par où peuvent entrer les molécules odorantes) : en forme de poil, de plaque, de pointe ou de simple creux sur la cuticule, elles ont des parois assez épaisses qui parfois exsudent un liquide visqueux aidant à la capture des molécules transportées par l’air. Elles agissent surtout par contact direct avec la source émettrice de l’odeur. Les sensilles à rôle olfactif prédominant sont des poils ou pointes aux parois fines et percées d’une multitude de pores : chaque sensille en possède jusqu’à 3000, chacun de 10 à 15 nanomètres d’ouverture ; une antenne de bombyx du mûrier porte 17 000 sensilles de diverses tailles, ce qui donne près de 45 millions de pores susceptibles d’intercepter des molécules pour un papillon ! Quelques molécules odorantes peuvent susciter une réponse nerveuse des sensilles frappées et moins de 100 molécules suffisent à orienter le comportement de l’insecte récepteur. Ces performances en disent long sur l’importance de l’olfaction dans la vie des insectes et tout particulièrement dans leur vie sexuelle avec les phéromones. Ce sera donc l’objet d’une future chronique.
Histoire des antennes


Autres chélicérates : Opilion avec 4 paires de pattes et une paire de pédipalpes (en avant) qui ne sont pas des antennes 
Scorpion : la pince ne correspond pas du tout à une antenne transformée
En tant qu’appendices articulés composés d’articles en cuticule, les antennes sont cantonnées au sein des seuls arthropodes ; pour autant, elles ne concernent pas tous les Arthropodes. A la base de cet immense groupe, on a une divergence majeure entre deux grandes lignées : d’un côté, les Chélicérates (Arachnides et apparentés) chez qui il n’y a pas d’antennes (ni de mandibules) ; de l’autre côté, les Mandibulates (dotés de mandibules) qui possèdent presque tous des antennes. D’ailleurs, on attribue aussi le nom d’Antennates aux Mandibulates. Au passage, si on vous demande comment on distingue un arachnide (araignée, scorpion, tique, acarien, …) d’un insecte, outre la présence de quatre paires de pattes (au lieu de trois chez les insectes qui sont des hexapodes), vous pouvez aussi donner comme critère distinctif l’absence d’antennes !
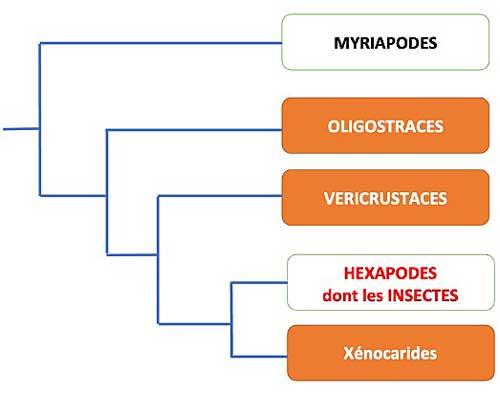
Le groupe des Antennates inclut les Myriapodes et plusieurs lignées divergentes de ceux que l’on regroupait autrefois sous le seul nom de Crustacés, aujourd’hui éclatés en plusieurs groupes : au sein de l’un d’eux (les Miracrustacés) émerge le groupe des Hexapodes comprenant les insectes. Au sein des Hexapodes, une des premières lignées basales non-insectes, les Protoures, en est dépourvue mais toutes les lignées d’insectes en possèdent au moins au stade adulte.

Les Ex-crustacés : Langoustine et ses très longues antennes 
Cloporte : il y a une seconde paire d’antennes mais très réduite 
Antennes d’un talitre (puce de mer)
Dans les lignées des ex-crustacés, il y a le plus souvent deux paires d’antennes (mais parfois la première est très réduite) comme on les voit très bien sur les langoustes par exemple ; la paire d’antennes des insectes correspond à la seconde paire des crustacés.

Les Myriapodes ont deux antennes : lithobie (Chilopode) 
Iule (Diplopode)
En dehors des Arthropodes, on trouve néanmoins une paire d’antennes chez des animaux très étranges à allure de vers segmentés à pattes multiples, les péripates (ou onychophores) des forêts tropicales ; leurs antennes sont bien segmentées mais pas du tout articulées ni faites de peau durcie ; il s’agit d’une convergence morphologique avec les antennes des Arthropodes. Enfin, en langage populaire, on attribue souvent le nom d’antennes aux appendices rétractiles et entièrement mous des escargots ou limaces : ce sont des tentacules et non pas des antennes !
NB Merci à Guillaume Lecointre (voir biblio ci-dessous) qui a attiré mon attention sur les onychophores.
Bibliographie
Classification phylogénétique du vivant. G. Lecointre. H. Le Guyader. Tome 2. 4ème ed. Ed. Belin. 2017
The insects. An outline of entomology. Third edition. P.J. Gullan ; P.S. Cranston. Ed. blackwell. 2005
Ant antennae: are they sites for magnetoreception? Jandira Ferreira de Oliveira et al. J. R. Soc. Interface (2010) 7, 143–152
Antennal hearing in insects -New findings, new questions. Björn Nadrowski et al. Hearing Research 273 (2011) 7e13
Antennal Mechanosensors Mediate Flight Control in Moths Sanjay P. Sane, et al. Science 315, 863 (2007)
